毛红椿混交树种选择及其幼龄林生长效应
2020-09-17黄红兰蔡军火
黄红兰,蔡军火,彭 欢,张 露
(1.江西环境工程职业学院,江西 赣州 341000;2.江西农业大学林学院,江西 南昌330045;3.江西特色林木资源培育与利用协同创新中心,江西 南昌330045)
高效混交林的营造是当代营林发展的一个重要方向,提倡阔叶树种和珍贵树种的多树种发展,依据树种的生态位、种间关系与立地条件等因素营建复层混交林[1-2]。我国成功的针阔混交林经验表明,灰木莲(MagnoliaglaucaBlume)、毛红椿[ToonaciliataRoem. var.pubescens(Franch.) Hand.-Mazz.]、火力楠(MicheliamacclureiDandy)、壳菜果(MytilarialaosensisLec)、顶果木(AcrocarpusfraxinifoliusWight ex Arn.)、降香黄檀(DalbergiaodoriferaT. Chen)、观光木[Micheliaodora(Chun) Nooteboom & B. L. Chen]、红椎(CastanopsishystrixMiq.)等优良阔叶用材树种,通过合理的混交模式,能消除针叶纯林的弊端、明显提高林分生产力、改善林地土壤肥力、增加物种多样性、提高立地质量,以及在促进林分的稳定、抑制病虫害、保持水土、森林防火和景观效果等方面均优于针叶纯林[3-15]。但是,由于混交林培育需要较长的周期,大多数混交林还处在试验性阶段,缺乏培育多树种混交的成熟经验[2],尤其对珍贵阔叶树种间的混交研究[8]。
毛红椿是我国珍贵的乡土用材树种,其人工林及生长特性的研究[16-17]取得初步成果,在福建等地与杉木[Cunninghamialanceolata(Lamb.) Hook.]、马尾松(PinusmassonianaLamb.)的混交效果理想,1 ∶4混交比例佳,林地土壤肥力得到改良,在针叶林阔叶化改造中得到较好的应用[18-20]。然而,我国毛红椿人工纯林、混交林处在幼龄阶段,培育技术体系尚不完善,与阔叶树种的混交尤其少。为此,从2011年起,在江西赣州等地营造毛红椿复层混交试验林,定期跟踪调查幼林生长,分析比较其在与观光木、火力楠、壳菜果和无患子(SapindussaponariaLinnaeus)等不同阔叶树种混交后的生长差异,探索适合南方林区瘠薄土壤山区的混交经营模式。
1 试验地概况
试验地位于江西省赣州市章贡区湖边镇湖边村下安组(114°54′38″E, 25°53′12″N),海拔110~125 m。该地属亚热带季风湿润气候,年平均气温19.4 ℃,无霜期286 d,年平均日照时间1 888.5 h,年平均降水量1 494.8 mm。试验林分位于5°~ 15°缓坡的下坡位,西南坡向,土壤为花岗岩发育的坡积母质黄红壤,土层厚度60 cm 以上,土壤肥力瘠薄。
试验地原林分为观光木×白花泡桐[Paulowniafortunei(Seem.) Hemsl.]、火力楠×中华杜英[Elaeocarpuschinensis(Gardn. et Chanp.) Hook. f. ex Benth]与壳菜果×苦楝(MeliaazedarachL.)的阔叶混交林,林龄均为5 a,株行距 2.5 m×3 m,块状分布,每种混交类型各3块,规格为(20~ 25) m×30 m,总面积约6 000 m2,林缘区域为灌丛、草地,面积约2 000 m2。原林分是基于改良土壤肥力而改造造林,改造措施如下:全部伐除针叶树,保留孤立的乡土树种苦楝、白花泡桐、黧蒴锥(CastanopsisfissaRehder et E. H. Wilson),对飞播的南岭黄檀(DalbergiaassamicaBenth.)、银合欢[Leucaenaleucocephala(Lam.) de Wit]实生优树实施间苗;栽植前种植穴施有机肥5 kg· 株-1。
2 研究方法
2.1 林分营造
2010年3月,对原林分进行结构改造,毛红椿分别与上述优势树种的阔叶混交林混交造林,带状混交,混交比例1 ∶4,株行距 2.5 m×3 m,于原混交林的林缘区域,新建无患子×毛红椿同龄混交林(混交比例4 ∶1)与毛红椿纯林(CK),按块状分布依不同林分间隔造林,分别设置3个重复,每块(10~15) m×(23~ 30) m,约300 m2,共6块,块状林间设置隔离带,带宽1.5~2 m,种植红花檵木(Loropetalumchinensevar. rubrum Yieh)灌丛。毛红椿、无患子裸根苗造林,苗木均为1年生的Ⅱ级以上实生苗木,株行距 2.5 m×3 m。造林当年,调查成活率,补植死亡苗木,林分的幼林抚育管理按常规方法。为方便统计,将林分按改造前的原林分与改造后林分予以区分,改造后林分有毛红椿纯林(CK)及无患子×毛红椿(SsT)、观光木×毛红椿(MoT)、火力楠×毛红椿(MmT)和壳菜果×毛红椿(MlT)混交改造林。
2.2 林分调查与土壤取样
在各块状林分中分别选设固定样地,至少包含1行毛红椿,规格10 m×20 m,共计15块,用于乔木层树种调查,并对样地每木调查,起测径阶5 cm。在每个固定样地的西南角选设5 m×5 m小样方1个,用于灌木层树种调查;在每个固定样地的四个角及其样地中心分别选设1 m×1 m小样方1个,用于草本层树种调查。2010年2月和2018年10月,对样地、样方进行林分因子调查与每木调查。
于2010、2019年的3月,连续晴天1周时取草本层固定小样方的表层0~ 20 cm土壤,然后分别将同一固定样地的5个小样方土样混均,按四分法取土用于土壤pH值及有机质、全氮、有效磷、有效钾含量的检测。2011—2018年10月初,每年观察各固定样地林分郁闭度(采用树冠投影法测算)和毛红椿的生长情况(实测胸径、树高)。
2.3 数据统计与分析
固定样地的毛红椿立木总蓄积为该样地毛红椿总株数与其单株材积的乘积,其单株材积按范建华[19]的公式[V平均单株=0.000 052 76D1.882 161H1.009 317,式中:D为胸径(cm);H为树高(m)]计算,其中,胸径、树高为对应样地的实测平均值。对应计算各类型林分的年平均生长量、连年生长量。固定样地其他树种(毛红椿之外)的立木蓄积采取二元材积法统计,样地林分立木总蓄积为毛红椿立木蓄积与其他树种的累计蓄积之和。固定样地林分的平均胸径、树高按不同树种的平均胸径、树高加权累加计算而得,某树种加权系数是该树种蓄积占林分总蓄积的百分比。固定样地林分定期总生长量分别为调查年度的平均胸径、平均树高和总蓄积,包括改造前的第5年间的总生长量和改造后第8年间的总生长量,其年平均生长量为对应调查年度的定期总生长量除以林龄计算所得,两次定期总生长量的增加值除以改造时间(8 a),为其改造林分的平均增长速率。此外,混交林的树种组成按各树种立木蓄积占所测林分总蓄积比例的十分法计核,当某树种立木蓄积占比为2%~5%,该树种前标记“+”号,而占比小于2%时,则标记“-号”。
数据统计分析用Excel和SPSS 20.0软件处理,方差分析的差异显著水平设置为a=0.05。
3 结果与分析
3.1 不同混交改造林分的结构差异
改造前,原林分的层次结构均较简单,以乔木层和草本层为主,灌木层树种少,零星散生桂花[Osmanthusfragrans(Thunb.) Loureiro.]或日本晚樱[Cerasusserrulatavar.lannesiana(Carri.) Makino],且长势极差。MoT原林分的乔木层优势树种是观光木、白花泡桐,主要伴生树种有红豆杉[Taxuschinensis(Pilger) Rehd.]、南岭黄檀,树种组成为9观+1泡-红;MmT原林分乔木层优势树种有火力楠、中华杜英,主要伴生树种有虎皮楠[Daphniphyllumoldhami(Hemsl.) Rosenth.]、南岭黄檀,树种组成为7火+2中+1虎;MlT原林分的乔木层优势树种有壳菜果、苦楝和白花泡桐,主要伴生树种有黧蒴椎、南岭黄檀和银合欢,树种组成为4壳+2苦+2白+1黧+1银。原林分草本层以阔叶杂草为主,种类不多,例如鸡眼草[Kummerowiastriata(Thunb.) Schindl.]、鸭跖草(CommelinacommunisL.) 和锈毛莓(RubusreflexusKer) 等,存在一定的种类差异。CK和MmT原林分为草丛,位于上述林分的林缘,种植麦冬[Ophiopogonjaponicus(L. f.) Ker-Gawl],白茅[Imperatacylindrica(L.) Beauv.]、马塘[Digitariasanguinalis(L. ) Scop.]和乌蔹莓[Cayratiajaponica(Thunb.) Gagnep.]侵入后,草本层优势种为麦冬和白茅,主要伴生种有马塘、乌蔹莓。
8 a林龄的混交改造林分,与原林分相比,乔木层树种组成发生较大的变化:SsT混交改造林分的树种组成是6无+4毛;MoT的是6观+3毛+1泡;MmT的为6火+3毛+中+虎+南;MlT的是6壳+1毛+1苦+1白+1黧。改造后林分的层次、结构趋向复杂,但灌木层与草本层的树种组成变化不大。
3.2 不同混交改造林分土壤质量的差异
原林分(含草地)的土壤常规检测结果显示,各林分间的各检测指标未见明显差异(表1) 。与建议标准相比,林地土壤的氮、磷养分元素均较严重缺失,表明原林分5 a间的生长,土壤肥力贫瘠情况未见明显转变。混交改造后的 8 a定期检测中,土壤有机质、全氮、有效磷及有效钾含量均比原林分明显提高,其中,MoT和MmT混交模式对林地土壤的改良效果最好,部分样地土壤的氮、磷含量接近建议标准的范围。可见,经复层混交的8 a间,不同混交林分的土壤质量均得到明显提高,达到较好的土壤改良效果。对改造前后林地土壤相同指标进行配对检验(表2) ,改造前后的差异极显著(t0.025=18.63~58.75,P=0.000) ,且混交改造后不同林分的土壤有机质、全氮含量间的差异达显著水平(表3)。

表1 不同林分改造前后的土壤成分Table 1 Soil composition of different forests before and after transformation

表2 林分改造前后的土壤成分配对检测Table 2 Paired detection of soil composition before and after forest transformation
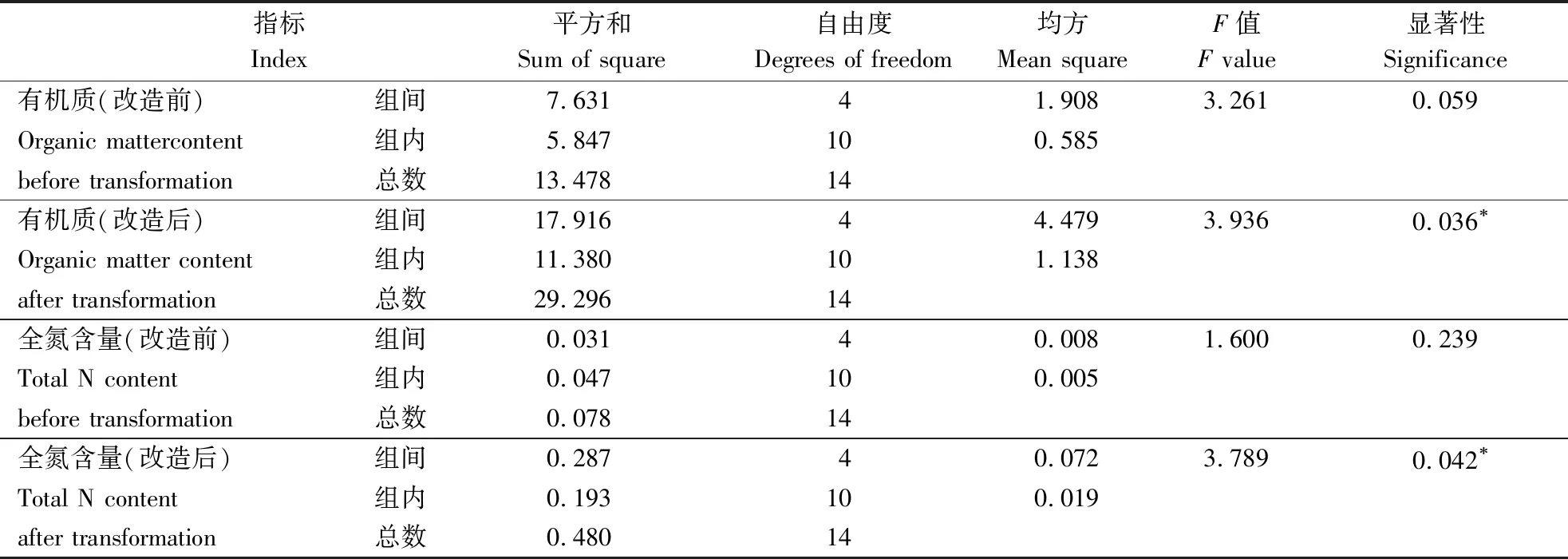
表 3 林分改造前后的土壤常规指标方差分析Table 3 Variance analysis of conventional indicators of forest soil before and after forest transformation
3.3 不同混交改造林分对毛红椿生长的影响
改造前与改造后林分生长变化见表4。改造前,林分立木的平均胸径、树高和林分总蓄积分别在3.8~4.2 cm、3.3~3.8 m和5.66~8.45 m3· hm-2之间,混交改造后,林分立木的平均胸径、树高和林分总蓄积分别在8.7~11.4 cm、7.8~11.3 m和27.43~81.75 m3· hm-2之间。表明经不同混交改造后,林分的生长得到提高。

表4 林分结构及其生长的变化Table 4 Changes in stand structure and growth
3.3.1 对胸径生长的影响 与不同阔叶树种混交改造后,随林龄的增长,毛红椿平均胸径呈“快-慢”的变化趋势。经方差分析与多重比较,各林分,4~8 a林龄毛红椿平均胸径均达显著差异,但大多未到生长高峰期(表5)。与CK相比,在MoT、MmT混交改造林分中,毛红椿于5~6 a林龄开始进入主林层,在6~7 a林龄出现了第1个小生长高峰期,各林龄毛红椿平均胸径均优于CK, 8 a林龄毛红椿平均胸径分别达12.1、12.7 cm,比CK分别增长了19.8%、25.7%,毛红椿胸径生长明显。 SsT、MlT混交改造林分中,各林龄毛红椿平均胸径均低于CK, 8 a林龄毛红椿平均胸径分别比CK低5.9%、30.7%,混交效果差,对毛红椿胸径生长有不同程度的抑制作用。其中,SsT混交改造林分中,毛红椿平均胸径经由 2~4 a较低的增速至后期趋于同时期CK增速的变化,但因其生长速度快于无患子,毛红椿基本保持在林分主林层,表明该混交改造模式对毛红椿生长存在抑制的可能性;MlT混交改造林分中,则先经 2~4 a保持与CK基本一致的增速生长后,5 a 林龄毛红椿退至亚林层, 5~8 a林龄毛红椿的胸径处于慢速增长阶段中,后期受壳菜果的严重抑制而滞长,胸径生长尤为缓慢。

表5 毛红椿胸径的年生长变化Table 5 Annual growth changes in average breast diameter of T. ciliata
3.3.2 对树高生长的影响 不同混交改造林分中毛红椿的树高生长及其年变化见表6。随林龄增加,毛红椿平均树高的增速基本均呈“快-慢”的变化趋势,于2~5 a间快速增长,随后增速不同程度地减慢,各林分,不同林龄的毛红椿树高均达显著差异。MoT、MmT混交改造林分中毛红椿的树高生长明显好于CK,在改造的第5~6年,毛红椿开始进入主林层,约于5.5、6 a林龄时,林分郁闭度高达0.8,随后生长缓慢, 8 a林龄毛红椿平均树高分别达12.7、12.5 m,比CK分别增长了22.1%、20.2%。而SsT、MlT混交改造林分中毛红椿的树高生长则均低于CK,混交受到抑制,随着林龄增长,差距进一步拉大。其中,SsT混交改造林分中,毛红椿树高生长在前期(4~6 a)低于CK同时期的增速,到后期反超 CK同时期的增速, 8 a林龄毛红椿平均树高达8.8 m, 比CK降低了15.4%;而MlT混交改造林分则抑制了毛红椿的树高生长,改造的3 a 林龄毛红椿的高生长优势被赶超而退居亚林层,至5 a 林龄毛红椿林分充分郁闭之后,其高生长几乎停止,8 a林龄毛红椿的平均树高最低,仅7.3 m。

表6 毛红椿平均树高的年生长变化Table 6 Annual growth changes in average stand height of T. ciliata
3.3.3 对材积生长的影响 不同混交改造林分中毛红椿的单株材积生长见表7。各林分,毛红椿的单株材积生长均处在增长期,但年生长增加趋势显现了较大的分异变化。在SsT混交改造林分中,毛红椿平均单株材积随林龄呈较平稳增速变化,但增速比CK慢, 8 a林龄的毛红椿单株材积比CK低 24.8%,生长效应表现为抑制可能型。 MlT混交改造林分中, 2~4 a 林龄毛红椿单株材积保持正常的生长表现,随后受壳菜果的上层冠层优势压制而极度滞涨,毛红椿的生长极差,8 a林龄的毛红椿平均单株材积比CK低64.9%,即表现为后期明显抑制型。MoT、MmT混交改造林分中,不同林龄的毛红椿平均单株材积生长量均优于CK,处在所有混交改造林分的前列,对毛红椿生长表现为促进效应,于4~6 a林龄快速增长,随后增速略减缓,8 a林龄毛红椿平均单株材积分别达0.074 9、0.080 7 m3,分别比CK提高了71.8%和85.1%,生长速度较快,年平均生长量达0.009 4 m3以上,比CK高71.8%以上。

表7 毛红椿平均单株材积的年生长变化Table 7 Annual growth changes of average volume per stand of T. ciliata
3.4 不同混交改造林分对郁闭度的影响
林分郁闭度是指森林中乔木树冠遮蔽地面的程度,是指示林分光照条件、林分密度及其评价林分质量的主要指标。原林分与混交改造林分郁闭度的年变化见表8。不同混交改造林分的原林分郁闭度程度比较一致, 在0.27~0.28之间;混交改造后,1~8 a林龄混交改造林分的郁闭度分别在0.11~0.72(CK)、0.10~0.56(SsT)、 0.34~0.89(MoT)、0.34~0.85(MmT)、0.32~0.89(MlT),均值在0.24~0.78之间。表明经不同混交改造后,林分的生长环境得到改善。
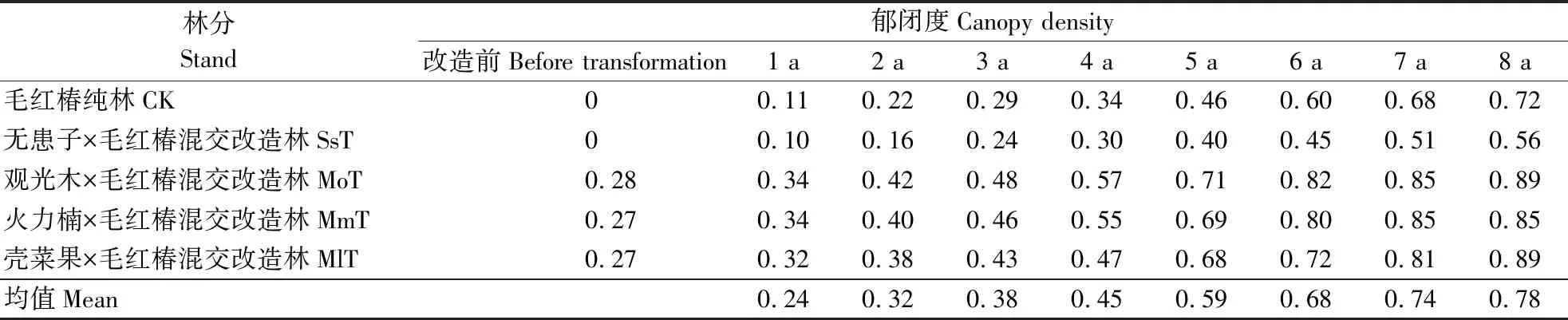
表8 改造前后不同林分的郁闭度Table 8 Canopy densities of different forests before and after forest transformation
试验林地土壤贫瘠,4 a林龄CK毛红椿胸径才达其起测径阶,明显低于林地质量好的同龄林分生长状况。1~3 a间,因数量少和基本处在其余树种的林冠下,可忽略其对混交林分的郁闭度贡献。假设混交树种的生长均稳定,与混交效果良好的前提下,4 a 林龄毛红椿开始进入林分主林层,之后的混交林郁闭度将会有较大的增幅。这一假设在MmT、MoT混交改造林分中有良好的验证。毛红椿早期耐阴,混交林林下的遮阴环境利于毛红椿幼龄林木早期快速生长,可促进其从更新层进入主林层。然而,在MlT混交改造林分中,壳菜果的前期遮阴下毛红椿速生优势正常,但当达 5 a及之后, 10 a林龄壳菜果的高生长完全突破毛红椿冠层层次占据到了主林层后,一直保持生长的优势,导致毛红椿的后续生长受到过度抑制。而SsT改造林中,无患子生长速度慢,毛红椿因此保持其主冠层的生长优势,但落后于CK的生长表现。这可能因无患子属落叶树种,且进入生殖初期,导致该林地枯落物少于CK,从而对土壤的改良效果较差。KOUTIKAetal[21]和GODINHOetal[22]研究也指出大量枯落物归还可改良土壤。可见,当选择与毛红椿种间关系表现良好的阔叶树种,调整合适的混交时间,与中郁闭度的林分混交,可形成促进毛红椿成为优势树种的良好生长效应的有利空间格局。
4 讨论与结论
毛红椿早期速生良好,可作为更新层的先锋树种,高优先生长特性使其占据林冠主林层,成为毛红椿分布区天然次生阔叶混交林的优势树种之一[23]。研究中,CK与不同混交改造林分之间的毛红椿胸径、树高生长差异显著,其中MmT、MoT混交改造林分中毛红椿生长良好,2~ 8 a林龄的毛红椿树高与胸径生长优于CK,且均处在生长表现的前列,随林龄增加基本呈“快-慢”的变化趋势,并表现为树高的生长节律优先(于5~6 a林龄林木达树高生长高峰),胸径生长有一定的滞后性(于6~7 a林龄林木达胸径生长高峰)。表明了MmT、MoT混交改造林分中毛红椿种间竞争的主导优势,这一优势同时有利于毛红椿材积增加。由范建华[19]核算的毛红椿材积公式可知,因胸径幕指数高于树高幕指数,随着毛红椿占据主林层,冠幅空间得以充分释放,胸径因而加大增速,不仅提高其单株材积,也有利于培育毛红椿大径级用材。CK及不同混交改造林分中的毛红椿单株材积处在增长速率期,这一现象分别与余林等[24]、徐海宁等[25]的研究结果一致。此外,前人研究认为毛红椿种内竞争促进树高生长[18-19],但本研究尚未得到验证,究其原因,可能是试验林地土壤贫瘠,在调查年度(2~ 8 a),8 a林龄CK郁闭度属中等水平,虽然种内竞争的压力大,出现了较显著的林木分化,但还未进入竞争激烈化的状态。
国外研究指出,混交林对土壤、根系及其物种多样性等方面随林龄、林分结构有不同程度的影响,有正面或负面的影响[26-28]。国内报道毛红椿与针叶树种混交林研究中,不利于毛红椿生长的不多。例如,前人研究杉木×毛红椿混交林,出现因杉木的侧方庇荫对毛红椿形成了胁迫,在竞争中促进了杉木、毛红椿的树高生长,但因杉木高生长优势明显,并不利于毛红椿占据主林层[18]。研究中MlT混交改造林分出现了物种间的树高交替竞争后,壳菜果最终进入主林层,出现不利于毛红椿逆转的局面。观察还发现,MlT混交改造林分会比CK与SsT混交改造林分更早进入首次抚育间伐期。至于抚育间伐,以及间伐后的林分生长表现及种间关系的后续变化,待继续调查研究。
