珠江口黄茅海底质沉积中介形类的生态类型及其分布特征
2020-09-10黄毅吴洁殷征欣关瑶倪锦锋
黄毅 吴洁 殷征欣 关瑶 倪锦锋


摘要:文章通过对黄茅海底质沉积样品中的介形类生态类型进行研究,探讨其空间分布特征。研究结果显示:研究区内出现的介形类可划分为4种生态类型,总体表现为以近岸广盐型分子占主导地位,优势度极高;浅海多盐-真盐型分子次之;而广海真盐型和淡水-微盐型分子丰度很低。根据黄茅海河口湾的水体条件,可大致将研究区划分为5种亚环境区,各区内的介形类生态类型组成也存在较大差别,表现出不同的空间分布特征。介形类分布受海水盐度控制最大,其次为水动力条件。在水体盐度小、水动力条件强的河流入海口及其影响区域,介形类丰度小,且生态类型单一;而在黄茅海河口湾南部海域,由于海水底层盐度全年在15‰以上,水动力条件适宜,介形类丰度总体较高,生态类型增多。
关键词:珠江口;黄茅海河口湾;介形类;生态类型;分布特征
Abstract:In this study,the ecological types of ostracod in bottom sediments of Huangmaohai estuary were analyzed to discuss their spatial distribution characteristics.The results showed that the ostracod could be divided into four ecological types in the study area.The dominant ecological type is the coastal euryhaline type,followed by the marine poly euhaline type.Besides,the abundance of the marine euhaline type and the freshwater slightly brackish type are very low.According to the water conditions of Huangmaohai estuary,the study area could be divided into 5 sub-environmental areas,and the composition of the ecological types of ostracod in each area is also quite different,showing the different spatial distribution characteristics.The results showed that the distribution of ostracod is mainly controlled by the salinity of seawater,followed by the hydrodynamic condition.In the estuaries of river with low salinity and strong hydrodynamic conditions,the abundance of ostracod is low and the ecological type is single.But the area in the southern estuary is different which shows the higher abundance and more ecological types of ostracod because of the bottom salinity of seawater is above 15‰ all year and the hydrodynamic conditions are suitable.
Key words:Pearl River Estuary,Huangmaohai Estuary,Ostracod,Ecological types,Distribution characteristics
微体动物介形类是生存环境范围较广的甲壳动物,常可见于海水、半咸水和淡水环境。介形类因其能敏感地反映水体盐度、温度、水动力条件、底质等环境参数[1],在地质学尤其是在分析海陆交互的河口地区环境上,具有重要的研究意义。因此,微体动物中除了有孔虫继续被充分研究和利用外,基于微体动物介形类的研究方法也成为探讨河口地带环境变化的手段之一。
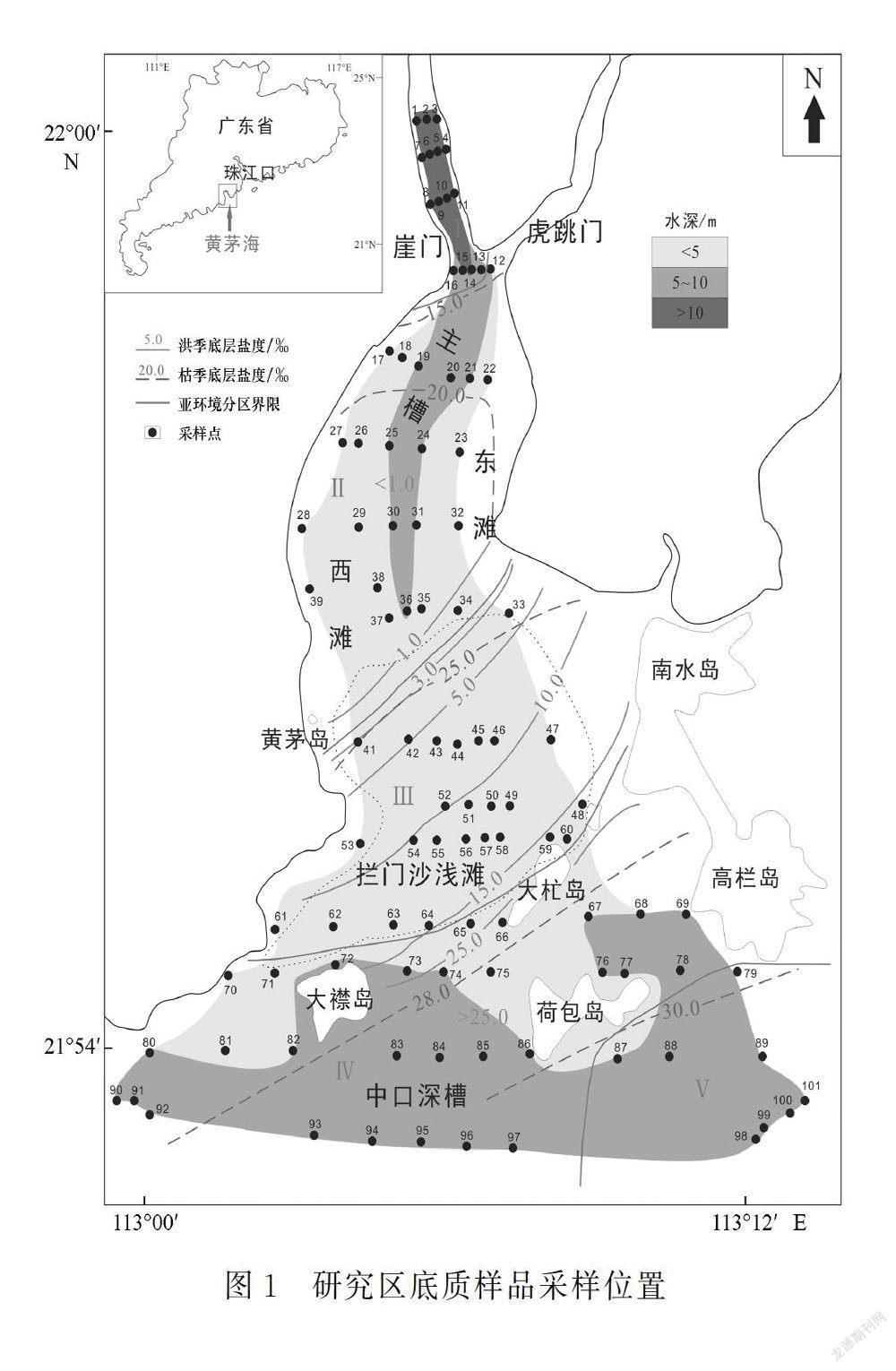
近年來对珠江口地区介形类微体动物的研究前人已有涉及[2-12],主要集中在伶仃洋海域表层沉积物或者三角洲钻孔样品。吴洁等[13]利用黄茅海有孔虫和介形类的去趋势对应分析,对控制微体动物分布的主要环境因素进行了初步研究。本研究在此基础上,对黄茅海的底质沉积样品中的介形类进行进一步分析,主要侧重于介形类生态类型及其空间分布特征。底质沉积样品取样站位如图1所示,具体取样和样品处理过程参见文献[13],盐度参见文献[14]。
1 研究区概况
珠江拥有世界上最复杂的河口系统,汇集西江、北江和东江等河网水系后,通过珠江口独特的8个口门入海[15]。珠江口发育有2个喇叭形河口湾,分别为东部的伶仃洋和西部的黄茅海,其中黄茅海湾顶端有虎跳门和崖门2个口门汇入,伶仃洋有4个口门(虎门、蕉门、洪奇沥、横门)汇入,而磨刀门和鸡啼门直接注入南海[16]。
黄茅海河口湾三面环山,呈NNW-SSE走向,海域面积约543 km2[15]。该海域内岛屿众多,通过岛屿峡口与南海相连[17],滩与槽交错,西滩、东滩、拦门沙浅滩的水深均在5 m以下,北部主槽、大襟岛和荷包岛之间的中口深槽的水深基本大于5 m,形成了黄茅海海域“三滩两槽”的水底地形格局[15,18]。
黄茅海河口湾属于潮流作用优势型河口,其潮汐为不规则半日潮性质,潮差由湾口向湾顶递减[19-22],并具有“东进西出”的近岸水流格局[23]。由于受到热带海洋性季风气候影响,河口地区的降雨量和径流量呈现明显的季节性,导致河口湾海域海水的盐度值也具有显著的季节性波动[24]。黄方等[14]研究显示,黄茅海河口湾水体盐度具有明显的季节性变化,枯水期海水上涌,湾顶口门处盐度可达15‰;丰水期受径流下泄影响,黄茅海湾内北部盐度较低。受黄茅海河口湾水流“东进西出”的影响,湾内的等盐度线整体呈NE-SW走向。研究发现,黄茅海河口湾北部主槽的表层沉积物中砂含量最高,其次为东口,中口深槽含砂量中等,拦门沙浅滩次之,河口湾东西两侧的浅滩砂含量最低,表明该地区不同水域水动力条件具有明显的差异[13]。
根据珠江口黄茅海海域盐度、水深、水动力环境等方面的差异,可大致将该海域划分为5个亚环境区(图1)。①Ⅰ区位于河口湾顶,河流入湾处,沿主槽水深10 m以上,主要受径流作用影响,水动力较强,枯季盐度小于15‰,洪季底层水盐度小于1‰。②Ⅱ区位于河口湾北部,拦门沙浅滩以北,枯季底层盐度15‰~25‰、洪季底层盐度小于3‰,受径流影响强于潮流作用,沿主槽水动力条件仍很强,水深5~10 m,东滩和西滩水深1~3 m。③Ⅲ区位于拦门沙浅滩海域,潮流上溯流与径流下泄流作用的交汇区,水动力条件相对较弱,水深3 m,两侧浅滩水深略浅,枯季底层盐度25‰~27‰,洪季底层盐度3‰~15‰。④Ⅳ区大致位于大襟岛—大杧岛一线以南,黄茅海湾口的南部海域,主要受潮流作用影响,枯季底层盐度26‰~30‰,洪季底层盐度15‰~25‰,水深5~10 m,近岛屿和陆地处水深较浅。⑤Ⅴ区位于黄茅海湾东口,水深通常大于10 m,潮水上溯流占优势,且水动力条件较强,枯季底层盐度大于30‰,洪季底层盐度大于25‰。
2 黄茅海介形类生态类型
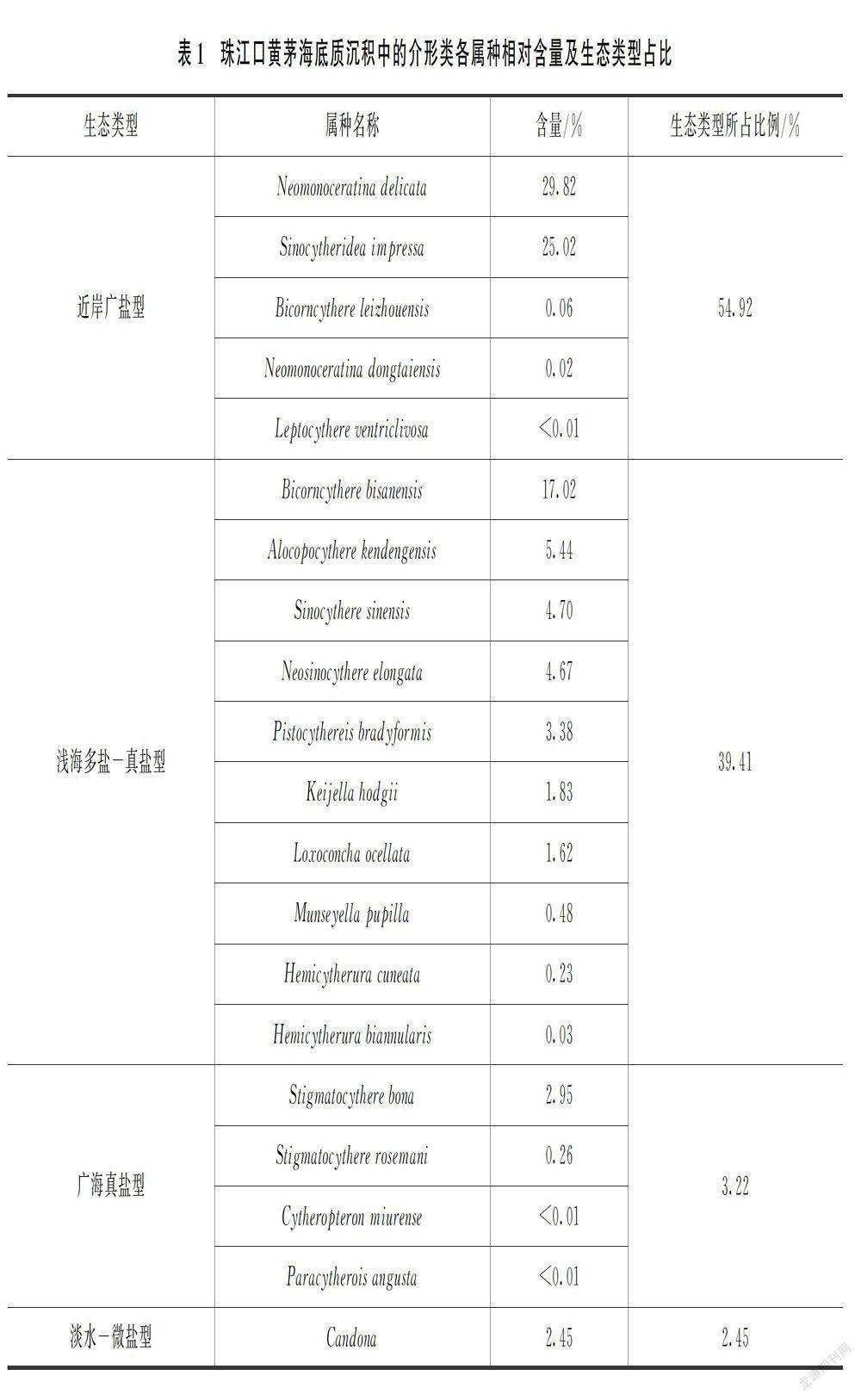
研究区底质沉积中共鉴定出介形类16属20种,其中Neomonoceratina delicata、Sinocytheridea impressa和Bicornucythere bisanensis这3个属种占比均大于17%,属于优势属种,其余属种优势度较低,各属种占比均小于6%,部分属种仅零星出现,各属种相对含量见表1。Liu等[25]曾对珠江口地区第四纪介形类的生态类型进行分类,本研究亦采用此分类方法进行划分和讨论。
受黄茅海河口湾多变环境的影响,该地区的介形类总体表现为以近岸广盐型分子占主导地位,优势度极高;浅海多盐-真盐型分子次之;而广海真盐型和淡水-微盐型分子丰度很低。
(1)近岸广盐型:该类型的介形类可适应盐度2‰至真盐,甚至高盐条件,盐度范围最广,多生存于小于20 m水深的沿岸水域,是黄茅海河口湾最为丰富的介形类。共发现5种,其壳体可占总数的54.92%。其中丰度最高的属种是Neomonoceratina delica和Sinocytheridea impressa。仅此2个种的壳体就占这一类型的99%以上。此外,还零星出现Bicorncythere leizhouensis、Neomonoceratina dongtaiensis、Leptocythere ventriclivosa等属种。
(2)浅海多盐-真盐型:这一类型介形类能适应相对较广的盐度范围,多分布于多盐至真盐条件、水深20~50 m的海域,仅少数有时可出现在沿岸水域中。浅海多盐-真盐型介形类在黄茅海河口湾中的丰度仅次于近岸广盐型,共发现10种,占介形类壳体总数的39.41%。主要属种包括Bicorncythere bisanensis、Alocopocythere kendengensis、Sinocythere sinensis、Neosinocythere elongata、Pistocythereis bradyformis、Keijella hodgii和Loxoconcha ocellata。其他丰度较低的属种有Munseyella pupilla、Hemicytherura cuneata和Hemicytherura biannularis等。
(3)广海真盐型:该介形类类型主要是在正常盐度的大陆架开阔海域,在河口地区其丰度和分异度通常都很低。广海真盐型的介形类在黄茅海河口湾发现4种,壳体仅占总数的3.22%。其中Stigmatocythere bona较为常见,其他属种仅零星出现,包括Stigmatocythere rosemani、Cytheropteron miurense和Paracytherois angusta等。
(4)淡水-微盐型:也可称为陆相型,多生活于淡水环境,偶在微盐水条件也可出现。该类型在黄茅海河口湾地区仅发现Candona属的1种分子,丰度低,其壳体仅占总数的2.45%。
3 讨论
珠江口表层沉积物中微体生物的空间分布可受水体盐度、水动力条件和水深的控制[11,13]。由于受潮汐作用和徑流作用的双重影响,不同位置的水体条件变化大,导致介形类丰度、生态类型组成变化大(图2和图3)。
(1)Ⅰ区:大多数样品中不含任何微体生物,仅少数样品见零星近岸广盐型介形类壳体,丰度小于50枚/100 g,且仅出现Sinocytheridea impressa和Neomonoceratina delicata 2个属种。由于该区域水动力很强[11],生物难以生存和保存,包括介形类。此外,受径流汇入的影响,该区域内底层水体盐度非常低(洪季底层水盐度小于1‰),也不利于介形类生存。
(2)Ⅱ区:介形类丰度一般在150枚/100 g以下,且壳体多为浅海多盐-真盐型小个体。受径流下泄作用外围的影响,该区域水动力仍然较强,且盐度较低(洪季底层盐度小于3‰),介形类仍较难生存。
(3)Ⅲ区:介形类丰度变化大(250~1 500枚/100 g),北部一般低于500枚/100 g,东侧受大杧岛影响,水动力较弱,丰度增加可达1 000枚/100 g以上。枯季受海洋影响显著,洪季仍受下泄淡水影响,盐度变化较大(3‰~25‰),适应力强的类型才能生存,区域内以浅海多盐-真盐型分子占主导,多数站位占50%,有的甚至达100%。浅海多盐-真盐型分子中Keijella hodgii比较丰富,有的样品中可达33%,该种虽属分布范围较广的浅海多盐-真盐型分子,但多分布于较浅水域。其次优势类型为近岸广盐型,总体占45%左右。此外还偶见海相分子。
(4)Ⅳ区:该区域盐度全年大于15‰,水动力条件中等,因此该区域是介形类最为丰富的区域,介形类丰度多为1 000~5 000枚/100 g,局部站位大于5 000枚/100 g。近岸广盐型多在50%以下;浅海多盐-真盐型分子减少,而广海真盐型分子比例增加。受黄茅海“东进西出”的影响,在黄茅海河口湾西口海域盐度整体较东口低,且由于离岸较近,近岸广盐型占优势。
(5)Ⅴ区:该区域内介形类丰度仅为中等,生态类型仍以近岸广盐型和浅海多盐-真盐型为主,但近岸广盐型相较其他水域略低,广海真盐型分子总体占比将近10%,部分站位可达34%。主要是因此处为黄茅海海湾东口,潮水常年从此处灌入黄茅海河口湾内,水动力条件较强,致使介形类丰度不高。同时,因该区域外侧为开阔海域,水体底层盐度常年大于28‰,广海真盐型分子得以发育。吴洁等[13]利用去趋势对应分析,认为盐度和沉积物含砂量(可代表水动力条件)是影响黄茅海地区介形类分布的主因。通过本次对介形类生态环境分布特征和分布环境的探讨,印证黄茅海河口湾介形类分布受海水盐度控制最大,其次为水动力条件。在水体盐度小、水动力条件强的河流入海口及其影响区域,介形类丰度小,且生态类型单一;而随着盐度由北往南升高,水动力条件逐渐适宜,介形类的丰度逐渐增加,生态类型更加丰富,在黄茅海河口湾南部海域,由于海水底层盐度全年在15‰以上,水动力条件适宜,介形类丰度和生态类型均达到最多。
4 结语
受黄茅海河口湾多变环境的影响,该地区底质沉积样品中出现的16属20种介形类可分为4种生态类型,总体表现为以近岸广盐型分子占主导地位,优势度极高;浅海多盐-真盐型分子次之;而广海真盐型和淡水-微盐型分子丰度很低。根据黄茅海口湾的水体条件可大致将研究区划分为5种亚环境区,各区内的介形类生态类型组成也存在较大差别,表现出不同的空间分布特征。研究显示介形类分布受海水盐度控制最大,其次为水动力条件。在水体盐度小、水动力条件强的河流入海口及其影响区域,介形类丰度小,且生态类型单一;而在黄茅海河口湾南部海域,由于海水底层盐度全年在15‰以上,水动力条件适宜,介形类丰度总体较高,生态类型增多。
参考文献
[1]WILLIAMS M A J,DUNKERLEY D L,DE DECKE P,et al.Quaternary environments[M].London:Arnold,1993.
[2]李淑鸾.珠江口底质中介形虫埋葬群的分布[J].热带海洋,1985,4(4):43-52.
[3]汪品先,闵秋宝,卞云华,等.河口有孔虫的搬运及其古环境意义[J].海洋地质与第四纪地质,1986,6(2):53-65.
[4]赵泉鸿,汪品先,张清兰.南海北部陆架底质中介形虫的分布[J].海洋学报,1986,82(5):590-602.
[5]赵泉鸿,汪品先.中国浅海现代介形虫的数量和属种分布[J].海洋与湖沼,1988,19(6):553-561.
[6]陈木宏.珠江崖门口沉积物中有孔虫分布的定量分析[A].中国科学院南海海洋研究所.南海海洋科学集刊第9集[M].北京:科学出版社,1989:29-37.
[7]罗宪林,李春初,牟崇键.伶仃洋有孔虫生物群和埋藏群的比较及河口沉积作用分析[J].海洋学报,2001,23(4):69-74.
[8]刘春莲,FRSICH F T,董艺辛,等.珠江三角洲PRD 05孔的高分辨率介形类记录与晚第四纪古环境重建[J].古地理学报,2008,10(3):313-322.
[9]李涛,向荣,李团结.珠江口表层沉积物底栖有孔虫分布及环境指示[J].海洋地质与第四纪地质,2011,31(6):91-98.
[10]吴洁,刘春莲,张素青,等.珠江三角洲东部晚第四纪微体动物群的古环境意义[J].中山大学学报(自然科学版),2014,53(6):45-52.
[11]WU J,LIU C,FRSICH F T,et al.Foraminifera as environmental indicators and quantitative salinity reconstructions in the Pearl River Estuary,southern China[J].The Journal of Foraminiferal Research,2015,45(3):205-219.
[12]吳月琴,刘春莲,杨小强,等.珠江三角洲中部全新世以来的微体动物群记录与古环境重建[J].海洋地质与第四纪地质,2019,39(2):31-43.
[13]吴洁,刘春莲,贾良文,等.珠江口黄茅海河口湾表层沉积物中有孔虫和介形虫的分布及其影响因素[J].古地理学报,2013,15(3):413-422.
[14]黄方,叶春池,温学良,等.黄茅海盐度特征及其盐水楔活动范围[J].海洋通报,1994,13(2):33-39.
[15]吴超羽.黄茅海河口小尺度动力结构及其沉积作用[J].中山大学学报(自然科学版),1995,34(2):86-94.
[16]李平日,乔彭年,郑洪汉,等.珠江三角洲一万年来环境演变[M].北京:海洋出版社,1991.
[17]应强,曹民雄,孔祥柏.黄茅海海域内泥沙淤积范围的确定[J].水科学进展,1997,8(1):48-53.
[18]贾良文,罗军,任杰.珠江口黄茅海拦门沙演变及成因分析[J].海洋学报,2012,34(5):120-127.
[19]吕颂辉,齐雨藻,钱宏林,等.南海港湾浮游植物与赤潮生物研究Ⅱ:广海湾[J].海洋通报,1993,12(2):57-62.
[20]张心凤,詹杰民.黄茅海波-流共同作用下的三维悬沙数值模拟[J].计算力学学报,2009,26(6):840-845.
[21]张心凤.黄茅海水域河床长期演变趋势预测[J].武汉大学学报(工学版),2014,47(5):591-598.
[22]吴创收,刘欢,武亚菊,等.黄茅海河口沿程异常潮差:I-理论模型研究[J].海洋科学进展,2010,25(4):436-444.
[23]杨雪舞,于红兵,孙宗勋,等.黄茅海河口湾现代动力地貌体系和冲淤过程分析[J].热带海洋,1997,16(1):49-59.
[24]DONG L,SU J,WONG L A,et al.Seasonal variation and dynamics of the Pearl River Plume[J].Continental Shelf Research,2004,24:1761-1777.
[25]LIU C,HUANG Y,WU J,et al.Miocene-Pliocene planktonic foraminiferal biostratigraphy of the Pearl River-Mouth Basin,northern South China Sea[J].Journal of Palaeogeography,2012,1:43-56.
