太湖蓝藻水华的年度情势预测方法探讨
2020-09-09朱广伟国超旋朱梦圆张运林秦伯强
朱广伟,施 坤,李 未,李 娜,邹 伟,国超旋,朱梦圆,许 海,张运林,秦伯强
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,太湖湖泊生态系统研究站,南京 210008)(2:南京师范大学虚拟地理环境教育部重点实验室,南京 210023)
在持续高投入治理10年之后,太湖蓝藻水华仍处于高发态势. 特别是2017年,太湖的蓝藻水华强度和水体总磷浓度双双反弹,给太湖蓝藻水华防控及周边城市饮用水安全保障带来了挑战[1]. 除了太湖之外,国内外多个类似大型湖库的蓝藻水华暴发强度都出现了高强度治理背景下的反复. 如巢湖近年来的治理投入大,蓝藻水华控制收效不佳,2012-2018年间,巢湖水质改善的效果变缓,虽然氨氮浓度继续下降,但2018年蓝藻水华的面积反而增加,水体总氮、总磷浓度也反弹增高[2]. 我国浙江省富春江水库,在“五水共治”水污染治理行动全面铺开、河道水体营养盐开始下降的情况下,却于2016年8月发生了较大规模的蓝藻水华状况[3]. 美国伊利湖自1960s实施了严格的营养盐控制措施,成功控制蓝藻水华数十年,但近年来蓝藻水华再次出现,2011年甚至暴发了历史上最大规模的蓝藻水华[4];2014年8月1日,因蓝藻水华引起的水厂出水藻毒素超标,伊利湖畔60万人口的城市Toledo中断供水2天[5]. 即使在水质优良的贝加尔湖,2016年7月26日也发生了数平方千米的长孢藻(Dolichospermum)水华[6]. Ho等通过遥感反演的方法评价了全球85个重要湖库过去30年的蓝藻水华变化趋势,发现大多数水体的蓝藻水华问题仍在加重[7]. 这些现象都表明,即使人类控制湖库富营养化的力度在加强,但是由于人类活动产生的污染负荷增加,损害湖库流域自净能力的土地扰动强度增加,气候变暖等有利于发生藻类水华的自然环境条件在恶化,湖泊和水库等重要水体的蓝藻水华风险仍将在较长时间内存在. 研发湖库蓝藻水华风险预测技术,提前对湖泊和水库的蓝藻水华发生强度及其引发的环境风险实施预测,指导管理部门充分应对,显得十分必要.
关于藻类水华灾害的预测预警技术,起步于20世纪末,近年来得到快速发展. 尤其是短期预测预报技术的发展迅速. 1999年,美国新泽西州Rutgers大学海洋与海岸带研究所的Schofield等提出了利用遥感技术与水体光学探头原位监测为信息支撑,采用简单推算模型的方法实施海洋赤潮短期预测的概念[8]. 随后,香港大学的Wong等在对近海赤潮短期预报的机理模型及条件阈值等方面进行了系统完善,并于2000-2003年在香港海湾实践了赤潮预测[9]. 美国海洋与大气局(NOAA)依托大尺度遥感监测信息,从2004年开始对墨西哥湾的腰鞭毛藻(Kareniabrevis)赤潮进行预测,从2008年起对缅因湾芬地亚历山大藻(Alexandriumfundyense)赤潮实施预测,2011年开始对伊利湖的微囊藻(Microcystis)水华实施预测[10]. Tian等则在深圳湾构建了完善的原位监测-遥感监测-水动力生态模型及基于GIS系统的结果展示海湾赤潮监测预测系统HMFS,实现了5 d的大面积海域赤潮短期预报[11]. 由于海湾环境变化和流场变化的复杂性,关于海湾的赤潮预测,目前都是一周以内的短期预报,未见有中长期预测的研究报道.
内陆水体的藻类水华预测技术方面,孔繁翔、秦伯强、李未等在太湖开展了大量的探索. 通过对太湖蓝藻水华不同生长阶段限制因子的辨识,孔繁翔等提出了以卫星遥感数据和湖面人工巡测为数据驱动、以未来1~3 d气象条件为预测情景条件的太湖蓝藻水华短期预测预警方法,并从2007年7月起在太湖开始应用[12]. 秦伯强、李未等则从蓝藻水华漂移堆积的水动力过程精准模拟入手,开发了基于三维水动力模型支撑的太湖蓝藻水华预测预警系统并长期实施业务化运行[13-15]. 叶麟和蔡庆华则基于逐日单点水质及环境参数的监测,开发了利用人工神经网络(ANN)预测未来7 d的大型水库支汊蓝藻水华风险预测方法[16]. 此后,大量关于湖库蓝藻水华短期预测的方法被报道. 这其中绝大多是基于模型算法方面的短期预测尝试和改进[17-21].
关于年际尺度、季度尺度的中长期蓝藻水华情势预报技术的报道很少. 与海湾相比,湖泊和水库生态系统更加封闭,环境条件更加稳定,具备在更长尺度上实施蓝藻水华强度预测的可行性. 孔繁翔等根据蓝藻水华形成的“四阶段假说”理论,基于2007年的蓝藻种源情况、影响蓝藻水华强度的水环境现状及2008年的水文和气候预测,对2008年的蓝藻水华开始时间、夏季强度及秋季结束时间等进行了定性预测[12]. 美国海洋与大气局的Stumpf等从2012年开始对伊利湖的蓝藻水华情势实施了提前一个月到提前一个季度的预测尝试[22-23]. 通过构建春季经由西伊利湖主要污染入流河道Maumee河的入湖有效磷负荷与夏季蓝藻水华强度之间的关系,建立了伊利湖夏季蓝藻水华强度的定量预测方法[23]. Obenour等在其基础上,通过贝叶斯层次模型进一步优化了预测模型的应用范围,将水华期藻类叶绿素a浓度与春季磷负荷的响应关系细分为3类,分类使用不同模型进行预测,提升了模型预测的精度[24]. 尽管该预测结果不尽理想,但是该预测的思路为进一步开发中长期蓝藻水华预测技术提供了借鉴.
太湖是国际上关于湖泊蓝藻水华情势及环境条件观测记录最完整的湖泊之一. 基于太湖湖泊生态系统研究站(简称“太湖站”)的观测记录数据库,回溯太湖蓝藻水华年度强度与环境因子之间的关系,有望建立切实可行的湖泊蓝藻水华季度/年度情势预测方法. 因此,本文通过对2005-2019年太湖蓝藻水华情势及相关水质、水文、气象关系的统计分析,试图构建利用早春气象、水文及营养盐条件与年度蓝藻水华强度之间的定量关系,探索一种能对全年蓝藻水华情势实施定量预判的方法,为太湖及类似的富营养湖库藻类水华防控提供理论支撑.
1 材料与方法
1.1 年度蓝藻水华强度估算
采用两种信息综合对太湖的年度蓝藻水华强度进行评估. 第一,太湖年度水体浮游植物叶绿素a人工监测的加权平均浓度. 这是一种能够定量反映全湖全年浮游植物生物量的指标. 本文采用太湖站每年2月、5月、8月、11月4次全太湖32个观测点采样(分上、中、下3层采样后混合水样)分析获得的水体浮游植物叶绿素a浓度,按照泰森多边形的方式获得各个点位的水量权重,加权计算出全年水体浮游植物叶绿素a浓度平均值,计作[Chl.a]LB,具体布点及加权平均值计算方法见文献[25]. 第二,采用每月多幅遥感影像获得的全年太湖蓝藻水华面积平均值,记作ABL. 这是一种定量精准反映全湖水华分布及其动态变化的指标. 本文采用的太湖蓝藻水华面积及其叶绿素a的遥感反演方法见文献[26],遥感影像数据采用MODIS aqua L-0数据,2005年1月1日-2019年12月31日的5000余景的逐日影像从NASA的Goddard Space Flight Center Web网站下载(oceancolor.gsfc.nasa.gov),经目视遴选,共获得太湖湖面无云可用的1522景影像实施水体叶绿素a浓度反演,反演的叶绿素a浓度记作[Chl.a]RS,其反演算法[27]为:
[Chl.a]RS=-1545.3- [(EXP(Rrc(645))-EXP(Rrc(859)))/
(EXP(Rrc(645))+EXP-(Rrc(859)))]+69.346
(1)
在本文中,[Chl.a]LB、[Chl.a]RS、ABL均可以在一定程度上反映当年的蓝藻水华强度. 为了更为客观地反映蓝藻水华的藻类生物量和水华面积特征,构建蓝藻水华强度指数BI(Bloom Intensity),定义BI为[Chl.a]LB与ABL的乘积与统计期间(2005-2019年)该值最大值的比值.
(2)
式中,i是具体某年份,而max则是2005-2019年的最大值,为2017年. 按此公式,2017年的BI值为100,其余年份与之相比. 对未来值而言,BI值允许大于100.
1.2 相关环境因子数据及其来源
根据施坤等对2003-2017年太湖蓝藻水华物候学的分析[26],温度、降雨、风速、营养盐等均为影响太湖蓝藻水华情势变化的重要环境条件. 从预测的操作性方面,我们分别从气象、水文、营养盐3个方面考虑影响年度太湖蓝藻水华强度的环境因子.
气象要素方面,从太湖湖泊生态系统观测站获得2004年1月1日-2019年12月31日太湖站气象观测场逐日降雨量数据. 由于前期的降雨量大小直接影响了外源引发的湖泊营养盐的积蓄程度,我们分3个时段进行分析,前一年降雨总量(RFYB,mm),前一年12月至当年3月降雨总量(RF12-3,mm),以及前一年9月至当年2月降雨总量(RF9-2,mm). 之所以强调12-3月的累积量,是因为传统上该时段是藻类生长的缓慢期,也是蓝藻快速增长的物质和能量积蓄期.
水文数据方面,采用了太湖站2004年1月1日-2019年12月31日逐日观测的水位(WL)数据及水下50 cm的水温(WT)数据. 其中水位方面,提取了前一年12月至当年3月的逐日水位平均值(WL12-3,m). 水温方面,也提取了前一年12月至当年3月的逐日水温平均值(WT12-3,℃). 此外,根据孔繁翔等观测提出的野外9℃以上太湖蓝藻开始复苏的报道[12],也尝试了积温统计:也即,将12-3月期间,每天的水温减去9℃,然后,将其中的正值部分进行累加,获得该时段的水温有效积温,记作AT12-3;同理,依据1月之后水温开始大于9℃起至3月底的逐日水温获得的有效积温记作ATs,依据2月和3月的逐日水温获得的有效积温记作AT2-3.
水质数据方面,根据前期的研究报道,氮和磷均能影响太湖蓝藻生物量的变化[28],因此,综合考虑了水体氮、磷浓度. 但由于水体总氮(TN)、总磷(TP)浓度很大程度上受水体藻类生物量反馈的影响[29],因此,还特别采集了北太湖具有代表性监测点位的溶解性总氮(DTN)和溶解性总磷(DTP)浓度表征水体营养盐因素,在太湖站逐月监测的北太湖14个监测点中,剔除THL00和THL06两个分别位于梅梁湾梁溪河口和直湖港河口的点位,将其余12个点位的TN、DTN、TP、DTP进行月值数学平均,然后,再将前一年12月至当年3月共计4个月的数学平均值求出,分别记作TN12-3、DTN12-3、TP12-3和DTP12-3.
1.3 数据统计处理
蓝藻水华强度与相关环境因子之间的定量关系分析,采用SPSS多元回归分析中的进入回归法依次分析. 各环境因子与蓝藻水华强度的关系,采用相关分析进行比选.
2 结果
2.1 2005-2019年太湖蓝藻水华强度变化
2005-2019年太湖水体[Chl.a]LB、ABL、[Chl.a]RS以及BI的变化如图1. 从图中可以看出,2017年所有指标出现峰值,年均蓝藻水华面积达到284.28 km2,是15年平均值(180.62 km2)的1.6倍;年均[Chl.a]LB达到40.8 μg/L,是15年平均值(19.6 μg/L)的2.1倍.
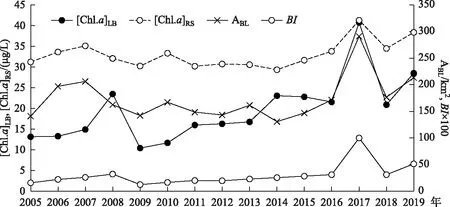
图1 2005-2019年太湖蓝藻水华强度指标变化Fig.1 Variation of annual cyanobacterial bloom intensity in Lake Taihu from 2005 to 2019
表征蓝藻水华年度强度的指标[Chl.a]LB和ABL在年度变化上并不完全一致. 水华面积ABL在2007年和2006年分别出现第2峰值和第3峰值,而反映藻类生物量的指标[Chl.a]LB则分别在2008年、2014年等形成与面积变化不同步的峰值. 2008年和2014年浮游植物叶绿素a浓度的反常增高,与同期藻类群落结构变化,特别是硅藻门生物量的明显增高有关[30]. [Chl.a]LB与[Chl.a]RS在变化趋势上基本一致,但遥感反演的值明显偏高,[Chl.a]LB与[Chl.a]RS多年均值分别为19.6和33.1 μg/L. 遥感值偏高是由于蓝藻水华物质的表层聚集现象所致,表层水体藻类生物量高于水柱平均值是合理的. 此外,二者的相关性非常好,相关系数达到0.701(P<0.004,n=15).
2.2 相关环境因子变化
2005-2019年与蓝藻水华情势密切关联的气象、水文、营养盐等相关指标见表1. 从表中可以看出,与BI显著相关的指标包括RFYB、RF9-2、WT12-3、AT12-3等. 这反映出水温、降雨等环境变化对年度太湖蓝藻水华强度具有较强的决定作用. 而冬、春季节的水体TN、DTN、TP、DTP等指标甚至与BI呈不显著的弱的负相关关系. 2005-2019年太湖冬季及早春(12-3月)逐日水温、水位、降雨量以及前一年的降雨总量变化如图2所示. 可以看出,就12-3月的平均水温而言,15年来的平均值为8.50℃,其中2007年、2008年、2017年和2019年的平均水温超过9℃,也即微囊藻能够开始生长的野外临界温度[12]. 最大值出现在2017年,为9.83℃,比平均值高16%. 第2高值出现在2007年,为9.73℃,第3高值出现在2019年,为9.45℃,第4高值出现在2008年,为9.38℃. 最低值出现在2012年,为7.40℃. 与图1比较能够看出,2007年和2017年的太湖蓝藻水华大暴发,都经历了冬季及早春的水温异常偏高,这与之前的分析结果一致[29]. 同时表明,在进行太湖这种营养盐本底较高的湖泊水华情势预测时,水温是最应关注的环境要素.
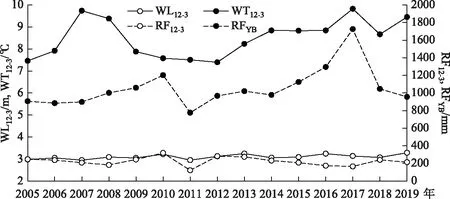
图2 2005-2019年每年12-3月太湖日均水温、日均水位、累积降雨量及前一年降雨总量的变化Fig.2 Variation of daily average water temperature, water level, accumulate rainfall during December and March,and rainfall of the ahead year in Lake Taihu from 2005 to 2019
与水温均值类似的参数积温,表现出与水温变化类似的趋势,也均与[Chl.a]LB等年度水华强度数据呈正相关(图3). 相比较而言,12-3月的逐日水温均值WT12-3与全年全湖平均浮游植物叶绿素a浓度[Chl.a]LB的相关系数明显强于积温. AT12-3尽管与[Chl.a]LB仍呈显著相关,但是相关系数明显小于WT12-3. 这表明,整个冬季的水温状况,而非仅仅是高于微囊藻复苏生长部分的积温,对全年藻类生物的影响更为重要.
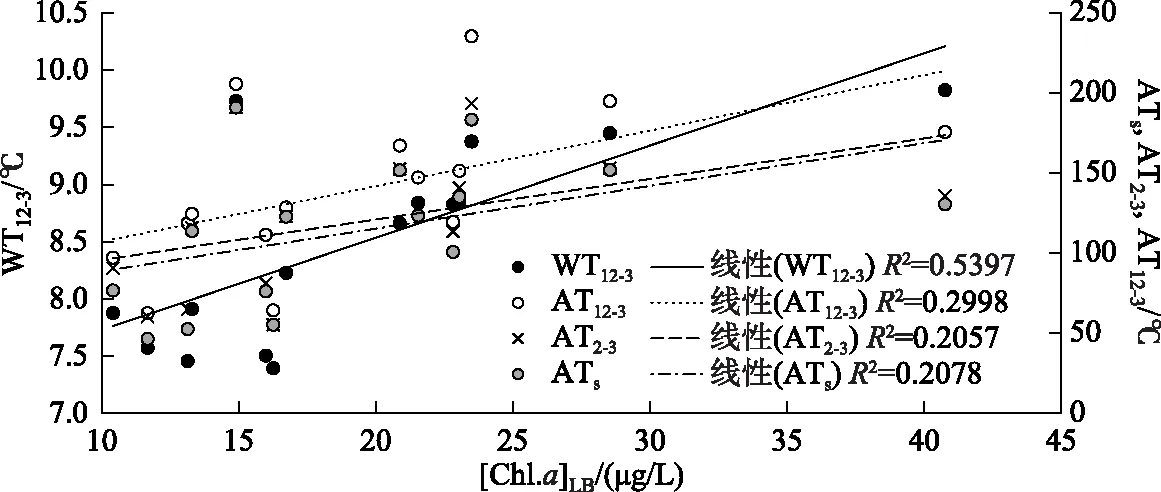
图3 冬、春季水温相关指标与[Chl.a]LB的相关性Fig.3 Relationship between [Chl.a]LB and water temperature parameters in winter and early spring
与之前的报道类似,北太湖水体TN、DTN浓度在2006年峰值以后,呈现不断下降的趋势[29]. 冬、春季是太湖水体TN浓度较高的季节,12-3月的北太湖均值在2006年为5.88 mg/L,2019年下降为2.95 mg/L,首次降至3 mg/L以下,下降幅度接近50%. DTN浓度的下降幅度更大,达到59%(表1). 然而,冬、春季水体TN浓度的下降与蓝藻水华强度变化和水体TP浓度变化很不一致,说明在当前的TN浓度范围内,氮的变化不是影响太湖藻类生物量的关键因素.
冬、春季(12-3月)北太湖TP浓度的最高值出现在2009年,为0.151 mg/L. 此后基本呈下降趋势,2017年为0.113 mg/L,与其他年份相比,水华异常严重的2017年年初水体TP浓度并不特别高. 这也从一个方面表明冬、春季水体磷浓度对夏季蓝藻水华情势影响也有限(表1). 2019年冬、春季北太湖TP浓度为0.102 mg/L,接近近15年来的最低值,与当年较为严重的蓝藻水华现象也不一致(表1).
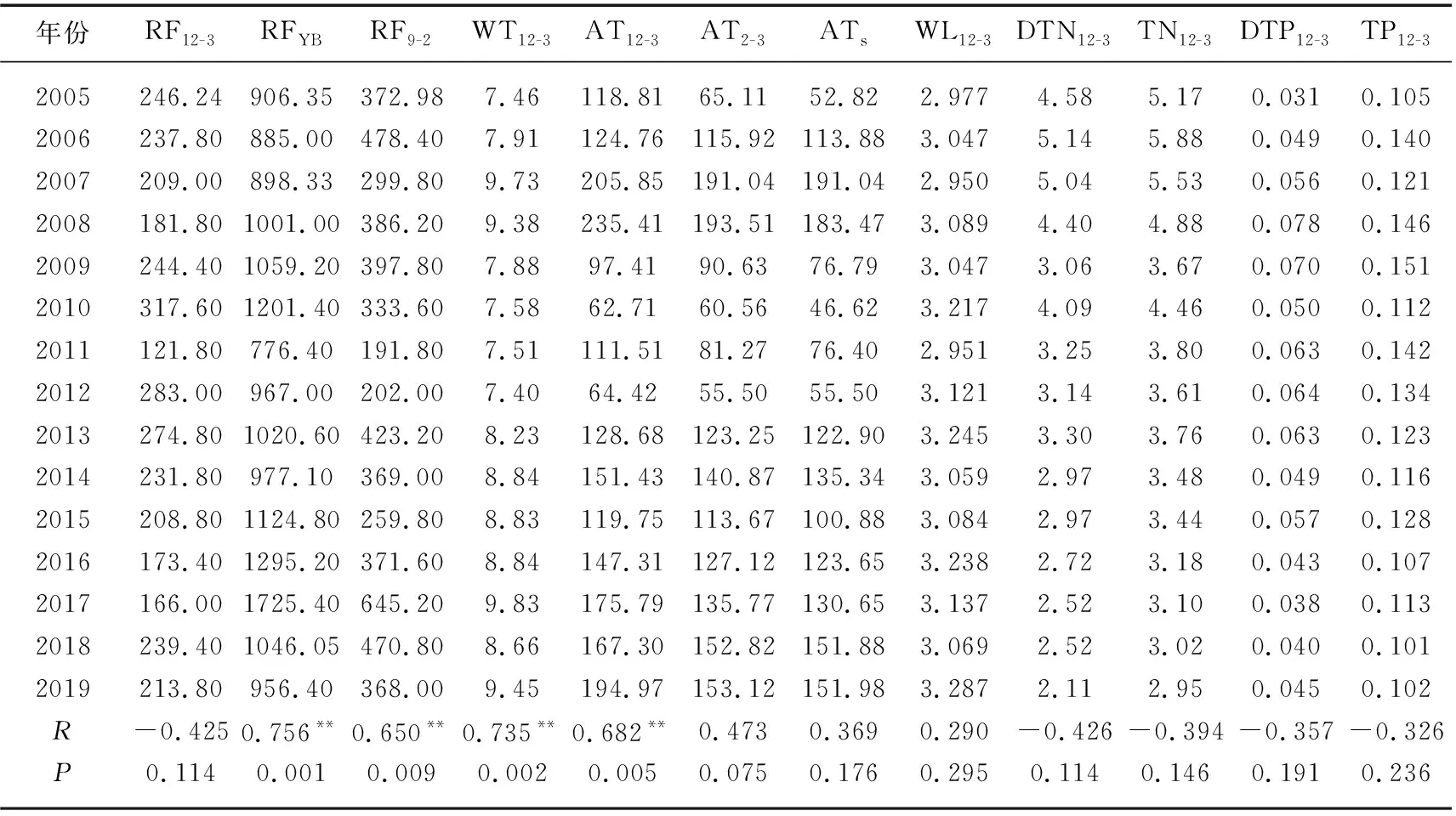
表1 2005-2019年太湖蓝藻水华情势相关环境因子及其与年度水华强度BI的相关性分析1)
2.3 预测模型构建
根据表1和图3,筛选了RFYB、WT12-3、DTN12-3、WL12-3、TP12-35个指标作为太湖蓝藻水华周年强度BI的预测因子,采用依次增加的方式进行逐步回归,比较各因子加入后对太湖年度水华强度的预测能力. 对数据进行正态分布分析发现,除RFYB之外的指标均符合正态分布. 为此,先对所有的数据取自然对数,然后再进行回归分析. 各回归方程及决定系数R2见表2.

表2 不同环境因子组合对太湖年度水华强度BI的预测关系式
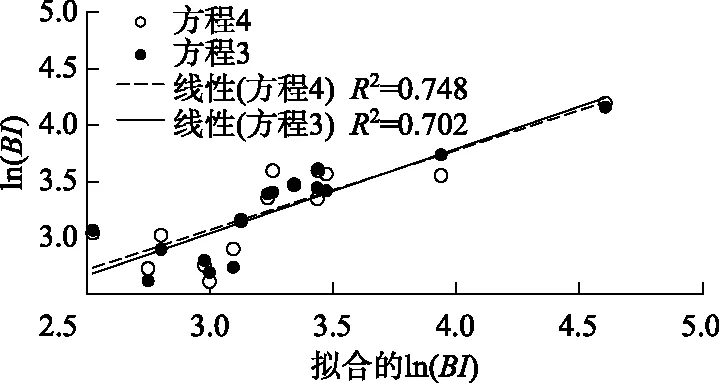
图4 太湖年度蓝藻水华强度实测值与方程拟合值的关系Fig.4 Relationship between calculated BI and simulated BI of Lake Taihu
对比表2中回归方程的决定系数,方程3和4具有明显的优势,特别是方程4,相关系数已经接近最大值,整个公式的显著性较高,入选参数简单易测. 而更多参数的方程,如方程5和6,对方程的精度提高有限,但增加了更多的参数需求,也增加了不确定性. 为此,采用方程4较为合理. 对比方程3和方程4拟合值与实际值可以看出,包含了水温、降雨量和DTN 3个环境参数的方程4拟合值与实测值相关性优于只有水温和降雨量2个参数的方程3(图4).
3 讨论
3.1 预测模型参数选择的合理性分析
越来越多的研究表明,水文气象因素是太湖等富营养化湖库蓝藻水华强度的主导因素. 影响湖泊浮游植物生长的主要因子包括光、温等气象条件,水动力、换水周期等水文条件,氮、磷、碳、铁、硅等营养盐条件,以及浮游动物、鱼类、大型水草等生态系统结构状况等. 王菁晗等统计了太湖、巢湖、滇池35年来的气象、水质因子的区别,发现适合藻类水华发生的时间长是滇池蓝藻水华较太湖、巢湖严重的原因之一[31]. 太湖的水体营养盐条件目前还明显高于能够限制浮游植物生长、蓝藻水华暴发的临界条件. 据许海等的原位浮游植物生长的营养盐添加实验判断,当水体溶解性磷浓度达到0.03 mg/L、溶解性氮浓度达到0.8 mg/L时,对藻类生物量增加的影响就不明显了,也即达到该浓度水体就能完全满足湖水中藻类快速增殖的需要[28]. 2005-2019年太湖全湖水体DTP浓度均值为0.053 mg/L,DTN浓度均值为3.45 mg/L. 其中2019年的年均值分别为0.045和2.110 mg/L,分别高出临界值50%和164%,因此,太湖全湖营养盐浓度并未低到能够对浮游植物生长产生限制的水平.
就空间上而言,2017年和2019年太湖年均水体DTN和DTP浓度如图5. 统计表明,2017年和2019年DTN浓度低于0.8 mg/L的面积分别为109和317 km2,也即藻类生长能够受到氮限制的区域增加了208 km2,约占太湖总面积的10%. 而2017年和2019年DTP浓度低于0.03 mg/L的区域面积分别为1703和1544 km2,2019年受到磷限制的水域面积反而减少了159 km2,凸显了太湖水体磷控制的复杂性. 从表1可知,太湖水体DTN浓度与全年藻类浮游植物浓度甚至呈不显著的弱的负相关,并不表明蓝藻水华强度的增加与氮的下降有关. 有两种可能的解释:(1)15年来太湖的治理带来的营养盐下降效应,被气温增加、雨量增加等对湖泊蓝藻水华生物量影响更大的环境因子的不利影响所抵消,出现了水体DTN浓度不断下降,但湖泊水体浮游植物叶绿素a浓度反而升高的结果,这种负相关,不代表二者具有因果关系;(2)水体氮浓度的明显下降,引起了浮游植物群落结构的变化,刺激了硅藻、丝状蓝藻占比增加,促进了水体浮游植物叶绿素a浓度的升高. 如果是这种机制,氮降低带来的叶绿素a浓度升高,虽然具有因果关系,但主要是非水华种藻类的增加. 这种可能性可以从近年来太湖直链硅藻等硅藻门生物量的快速增加得到支持[30]. 初步判断,可能是两种机制共同导致了太湖目前DTN浓度与水体叶绿素a浓度的负相关关系. 比较几个营养盐参数发现,15年来太湖冬春季DTN12-3浓度与[Chl.a]LB之间统计关系最好,因此,可以在蓝藻水华强度的预测中将DTN12-3列入作为营养盐要素代表.
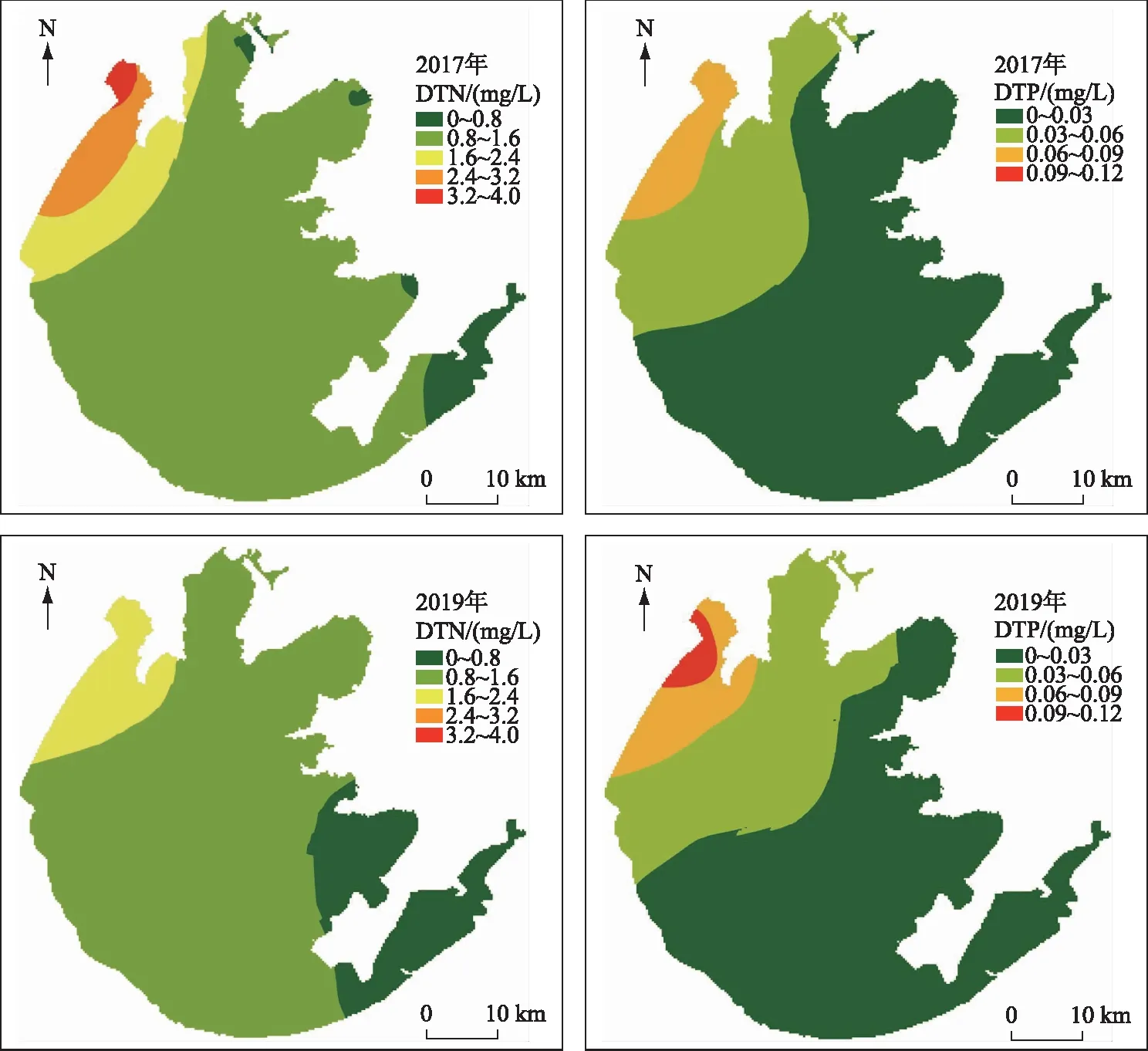
图5 2017年与2019年太湖年均DTN和DTP空间插值对比Fig.5 Comparison of spatial DTN and DTP distribution between 2017 and 2019 in Lake Taihu
使用温度和降水指标作为太湖年度蓝藻水华情势预测的关键参数既符合太湖实际,也具有合理性. 近年来,变暖、极端天气增多等气候变化与人类活动的影响交织,使得全球湖泊、水库等淡水水体蓝藻水华问题在加重. Huisman等系统分析了近年来气候变化趋势有利于蓝藻水华发生的机制,包括:(1)气温升高、热分层加剧,更有利于喜欢表层生长的蓝藻生长和细胞集聚,且下层缺氧几率增大带来的内源营养盐释放增加;(2)大气CO2分压增高,增加水体CO2供给,有利于水体浮游植物生长;(3)暴雨等极端气象事件增加带来的流域地表冲刷作用加剧,湖泊外源营养盐负荷增加等[32]. Huisman提出的气候变化有利于蓝藻水华的几种机制,近年来在太湖都有所表现. 施坤等通过太湖2003-2017年蓝藻水华物候学分析发现,气候变暖等导致近年来太湖蓝藻水华发生的物候学条件提前了近1个月[26]. 邓建明等研究发现,气温升高显著影响了太湖浮游植物群落结构的演替,气温、水温与微囊藻生物量的变化呈显著正相关[33]. 近年来全球普遍发生的风速下降现象在太湖流域同样存在,而风速的下降有利于易于上浮集聚的蓝藻细胞快速增殖和优势形成,叠加太湖近年来的水位升高,下层水体扰动减弱,又促进了太湖内源释放,共同加剧了太湖的蓝藻水华问题[34].
冬、春季水温的变化对微囊藻为主的蓝藻冬季种源、春季生物量基础有着重要的影响. 因此,水温成为预测年度蓝藻水华情势的重要考量要素是合理的. 平均温度显示出比有效积温更好的蓝藻水华强度拟合关系,说明太湖的蓝藻生长、蓝藻水华强度不仅受9℃以上的水温高低的影响,更低的温度对藻类种源的存量基础也有影响. 其原因可能有两点:(1)太湖的微囊藻在9℃以下死亡或生物量下降是个缓慢的过程,甚至成团之后的微囊藻能在9℃以下长期存在. 马健荣等发现太湖冬季蓝藻水华还能存在,表明短期低温下微囊藻依然存活、微囊藻水华尚可存在[35]. 此外,如果低于9℃时微囊藻细胞能够部分被冻死,抑或有助于食物链牧食降低微囊藻种源,那么,温度低于9℃也的确依然影响整个冬季的蓝藻生物量基础. (2)其他藻类的生长状况会间接影响夏季微囊藻的生长. 比如,冬季太湖的鱼腥藻大量存在,其生物量对后期微囊藻的演替具有影响. 此外,冬季硅藻生物量偏高,可能有助于营养盐在水相的累积,也有利于春、夏季微囊藻生长. 因此,选择WT12-3而非有效积温作为温度预测变量,对太湖蓝藻水华年度强度的预测更为合理.
降雨量高低对太湖蓝藻水华形成和发展的影响是多方位的. 太湖的降雨量不但代表了太湖的年度换水程度,还在很大程度上反映了外源综合污染的负荷量. 翟淑华等通过水量平衡及物量平衡的方法,估算了太湖2012年、2015-2016年的磷负荷特征,指出入湖水量大小是影响太湖磷收支的主要因素[36-37]. 朱伟等进一步分析认为2016年太湖流域特大洪水期间的洪峰携带入湖及太湖的高滞留特点对太湖磷收支影响很大[38]. 陈洁等分析了2017年3月1日-2018年2月28日间太湖主要营养盐入湖河道大浦河(陈东港)水体氮、磷浓度与降雨过程之间的关系[39],发现河道入湖水量是表征磷负荷的重要指标,河道降雨过程对入湖河道水体氮、磷浓度影响不大,大雨、中雨、小雨和无雨期间河道水体总氮浓度分别为3.00、3.34、3.55和3.37 mg/L,总磷浓度分别为0.228、0.258、0.219和0.225 mg/L,不同雨强之间统计上无差异,降雨量、入湖水量很大程度上表征了河道入湖的氮、磷负荷. 此外,降雨量大小还影响了太湖湖体的营养盐等物质的空间输移,间接影响蓝藻适宜生长区的时空动态变化. 比如2016年的暴雨增加了太湖高磷浓度水域的空间扩张[38],形成了明显的自西北向东南湖区推移趋势,增加了蓝藻水华易发区的面积,从而影响蓝藻水华情势. 因此,降雨量大小综合反映了水文和营养盐两方面对蓝藻水华情势的影响,纳入预测模型具有合理性. 由于秋、冬季降雨量不大,偶然性很大,因此利用前一年总降雨量RFYB对年度蓝藻水华强度的预测精度反而明显高于秋、冬季RF9-2或冬、春季RF12-3.
3.2 预测模型存在的不足
缺乏对未来气象情景预测信息的综合考虑是本预测模型的不足. 本预测模型主要从影响藻类生长的现有环境条件基础的角度开展未来藻情预测. 影响藻情变化的,除了现有条件基础,还与未来春、夏季蓝藻水华暴发期间的水文、气象、营养盐条件密切关联. 因此,对未来春、夏季气象水文情势的预测信息未能体现,是本预测模型的不足. 目前对气候预测的信息渠道比较多,有一些非常专业的机构对外发布报告,可将其纳入,将有助于对未来蓝藻水华情势的判断.
缺乏对藻类群落结构变化的深入分析会影响本预测模型的精度. 本预测也未考虑藻类群落结构信息的影响,泛泛采用在一定程度上反映总藻类生物量的叶绿素a浓度年均值表征蓝藻水华强度,在藻情评估方法上尚有一定的不足. 太湖藻类群落结构的季节变化较大,空间差异性也很大[30],因此,仅仅采用藻类叶绿素a浓度表征有害蓝藻生物量、微囊藻生物量存在一定的不足. 目前,由于藻类群落结构数据的监测、可比性仍待提高,优质数据的获取存在难度,纳入模型之后可能会降低预测工作的时效性. 建议在今后模型的优化中,当藻类群落结构快速检测技术成熟时[40],将其纳入预测模型中.
致谢:太湖湖泊生态系统研究站钱荣树、黄建明、韦金权、夏丽萍等及时提供了2019年监测数据,在此表示感谢.
