有花植物与传粉媒介溯源
2020-08-28

水生被子植物辽宁古果是迄今为止最早发现的被子植物化石之一
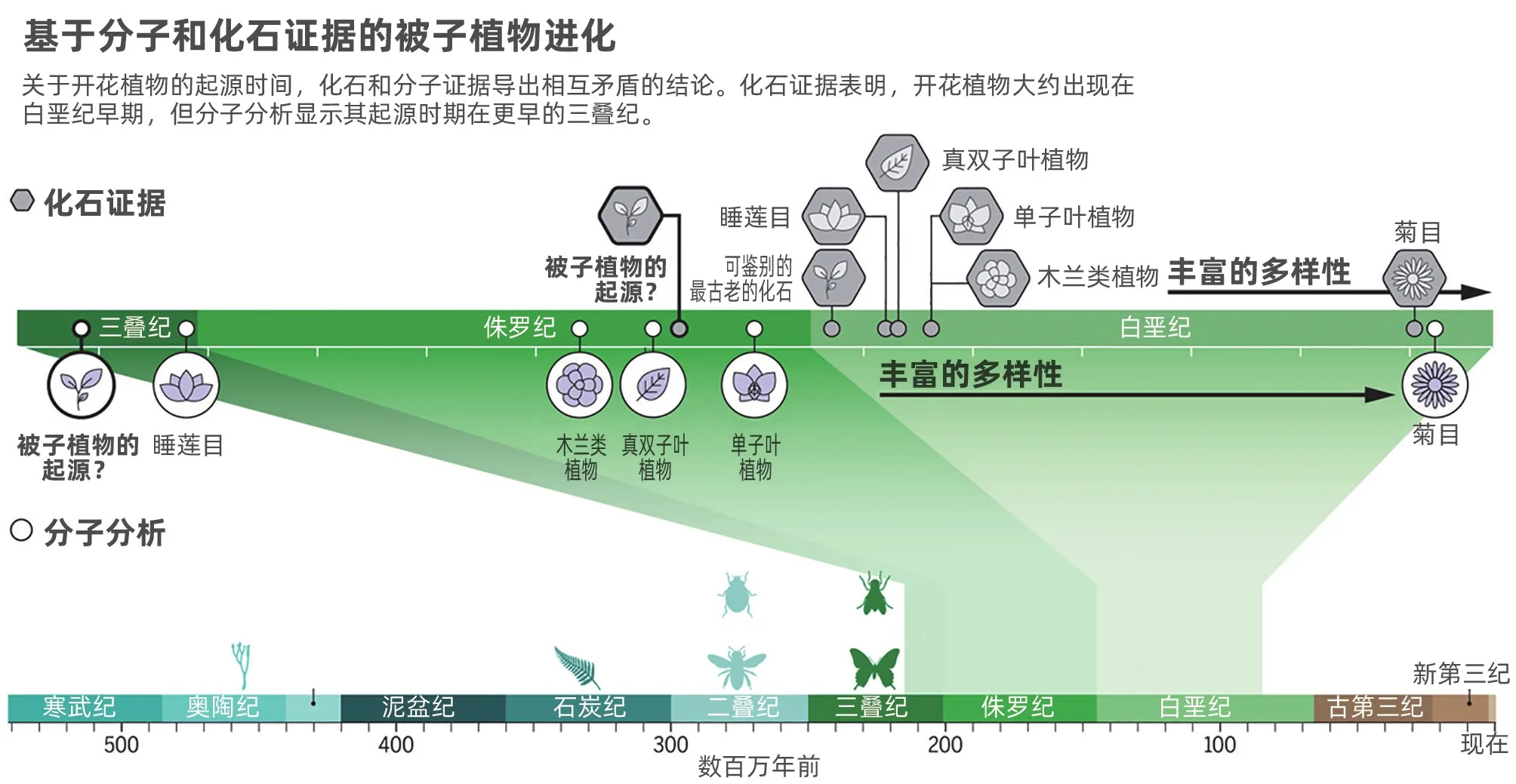
一个多世纪以来,人们一直对开花植物(被子植物)惊人的增长速度和早期的多样性感到着迷。达尔文将被子植物看似爆发性的多样性描述为“讨厌的谜团”,而关于促使被子植物物种形成的起源与过程的争论一直在继续。传统上,被子植物起源的年代测定是研究植物化石记录的古植物学家的特权,然而随着DNA测序技术愈加成熟,分子年代测定法开始走上舞台。许多被子植物化石可以追溯到早白垩纪(大约1.35亿年前),这使得古植物学家可以推断出它们起源于哪个时代。现在人们越来越多地认识到被子植物可能比最古老的化石还要古老,但究竟有多古老仍颇有争议。被子植物的起源是弄清楚传粉媒介——尤其是蜜蜂、蝴蝶、飞蛾和苍蝇等昆虫——起源和进化的关键。
最近的报告强调了分子和古生物学时间尺度上的差异,并对被子植物多样化的时间得出了相互矛盾的结论。一项基于85%的现存开花植物科的2 881个叶绿体基因组的基因序列,利用62个化石进行时间校准的研究,将被子植物的起源时间认定在晚三叠纪,即两亿多年前。这比最早被认证的被子植物化石的形成时间还早了约7 000万年(大致相当于侏罗纪时期)。这项研究进一步表明,主要的放射状分布(物种多样化)发生在侏罗纪晚期和白垩纪早期,距今约1亿~1.65亿年。相比之下,对古植物学证据的综述反驳了白垩纪之前即出现了规模多样化这一观点,而是认为只有一些特定的演化支(如睡莲)可能起源于侏罗纪晚期。不同类型化石和形态特征的顺序出现,提示主要的放射状分布不太可能出现在更早的时期。虽然被子植物出现在白垩纪早期这一概念似乎很难与该时期观察到的形态多样性的迅速增加相一致,但是如果在白垩纪放射状分布迅速发生,也不是不可能的。
古生物学记录和分子分析各有优缺点。化石的优势在于,它们可以提供关于过去的形态、功能和演化支丰富程度的信息,并间接提供了关于物种形成和灭绝的信息。当化石包含过渡结构或已不存在的特征组合时,它们就特别有用,这可以提供帮助重建进化事件过程的深刻例子。然而,对化石的解读可能具有主观性和争议性,因为这些植物的重要特征可能没有被保存下来,而且往往必须从二维压缩化石中推断出来。
证据的缺失正是因为没有证据来证明化石的缺失,而且我们知道,化石记录可能是不完整的或片面的,因为有一些类群可能不容易形成化石。例如,尽管花粉是一个例外——因为它通常能在更极端的条件下生存——但特定的生态环境或栖息地将影响整株植物化石化的可能性。除此之外,将化石定位在某一特定时期,需要对发现化石的地层进行准确的年代测定,虽然这一因素造成的误差通常很小,但还是会出现问题。重要的是要记住,在起源时间和最早可识别的化石之间可能有相当大的时间上的滞后,因为化石通常出现在某一生物分类单元已经存在一段时间并且出现的频率相对较高的时候,这种现象被称为模糊效应。
被子植物的特征主要表现在它们与传粉动物,尤其是与昆虫的关系。与植物一样,昆虫的多样性也是一个充满不确定性的领域。访花昆虫中几个重要目,例如鞘翅目、双翅目、膜翅目和鳞翅目,起源于二叠纪或三叠纪(2亿~3亿年前),在白垩纪有着明显的多样化时期,这与被子植物主要的放射状分布年代相吻合,故常被提及。然而,访花昆虫起源时间的认定仍具争议。例如,根据化石证据提出了针对鳞翅目(蝴蝶和飞蛾)的放射状分布发生于晚三叠纪这样的说法,但最近一项针对几乎覆盖所有鳞翅目超科转录组的研究将其起源追溯到了更早时期的石炭纪(约3亿年前)。
关于被子植物的起源时间仍存在争议。如果被子植物出现在侏罗纪之前,这对理解昆虫授粉的进化方式将具有深远的影响。毫无疑问,昆虫授粉加速了被子植物的辐射状分布。然而,根据最新的发现,究竟哪个因素引发了何种进化事件变得更加错综复杂。长期以来,人们认为早期分化的无花种子植物(裸子植物)的风媒传粉为被子植物的动物媒传粉所取代,并认为这种由风媒传粉向动物媒传粉的转变导致了被子植物的多样化,但这种认识似乎过于简单化了。
许多现已灭绝的裸子植物(如本内苏铁目)是虫媒传粉的,被子植物可能是直接从虫媒传粉的裸子植物进化而来,也可能是从风媒传粉的裸子植物进化而来,它们通过选择为同一群落中的裸子植物服务的昆虫进行进化。相反,如果被子植物真的起源于早期三叠纪,一些裸子植物可能会从早期的被子植物中选择昆虫作为传粉者。然而,后一过程在被子植物的进化过程中似乎不太重要,因为即使这一进化过程发生在更早的时期,被子植物也不是侏罗纪时期的优势植物群。相比之下,在晚三叠纪至侏罗纪的植物群中,本内苏铁目及其他早期种子植物具有生态优势,说明被子植物向虫媒传粉的转变是从这些裸子植物类群开始的。这些可能性比预想的从原始风媒传粉到高级虫媒传粉的标准情景要复杂得多,它们暗示着更加丰富的生态环境。物种之间的相互作用比以前所认识的也更加复杂,包括现在作为传粉者不太重要的昆虫群,例如长翅目(蝎蛉科)。
开花植物起源的时间也为它们最显著的特征——花的进化提供了最低年代。在许多现存的裸子植物(如苏铁类植物、麻黄属、买麻藤属)中,虫媒传粉的进化主要是通过气味而不是视觉吸引。已经灭绝的裸子植物可能也是如此,但因为气味不会变成化石,我们永远都不可能知道真相了。然而,如果这些已经灭绝的裸子植物的生殖结构功能与它们现存的近亲相似,也就是说以气味为主导,那么在被子植物中视觉线索对吸引传粉者的重要性就会与日俱增,这可能是被子植物进化成功的决定性特征之一。此外,花的结构起源早于一些特殊的访花昆虫目,花的特征也因此促进了这些大的昆虫群体的特征进化。
在某些系统中,有明确的例子表明特定的花和传粉者形态特征的共同进化,比如花管长度和传粉昆虫舌的长度。那么颜色和气味等花卉特征是怎么进化的呢?例如,也许花的颜色和气味是为了适应传粉者的视觉和嗅觉而进化的,反之亦然。或者,信号的产生与其探测方式可能是同步进化的。昆虫颜色视觉的基本原则,如拥有三种光感受器(紫外光、蓝光、绿光),其产生时间似乎早于花的产生时间,不管是出现在三叠纪还是更晚的时候。因为色觉在一些关键行为——比如探测潜在的配偶和捕食者,以及寻找产卵地点中也起作用,所以色觉的进化不太可能受到花色的驱动。
在一类由圣甲虫授粉的植物中也发现了相似的嗅觉产生的起源,与植物产生气味相比,授粉者接受气味的时间比植物产生气味信号的时间早。然而,嗅觉或色觉的行为学方面——比如决定了不同昆虫类群的觅食行为的先天性颜色偏好——可能进化得较晚,以响应花的信号。所有这些都取决于开花植物进化的时间,以及引发了虫媒授粉的进化事件的顺序。例如,如果虫媒授粉的裸子植物起源时间早于被子植物,那么追溯这些视觉和嗅觉特征的起源,就有可能追溯到曾经统治过陆地植物区系的早已灭绝的植物分支。
未来的古生物学探索一定会发掘出更多化石,并且互补测序方法和更复杂的进化模型的使用,将有助于减轻植物中广泛存在的多倍体所造成的限制,这种限制经常妨碍对核基因的分析。达尔文关于有花植物进化和放射状分布的时间疑问能否得到解答仍然是个谜,但显然这个问题及其对理解虫媒传粉的生态学意义是很复杂的。
资料来源 Science
