两种硫脂酶对重组菌游离脂肪酸合成能力的影响
2020-08-27侯艳玢闫依狄孟鑫
侯艳玢,闫依狄,孟鑫
(锦州医科大学食品科学与工程学院,辽宁锦州 121000)
脂肪酸作为一种重要的化合物,广泛应用于食品、医药、保健品等诸多方面,目前主要以动、植物油脂为原料生产[1],但存在油脂产量低、生产成本高等缺陷,限制了大规模工业化生产。近年来,已有多种微生物用于功能油脂的生产[2-4]。然而,微生物细胞内总脂需采用传统破壁、离心工艺获得,若脂肪酸能以游离形式分泌到细胞外,可减少复杂的分离工艺。已有报道在E. coli中表达植物或内源硫脂酶基因,可获得游离脂肪酸[5-7]。植物硫脂酶基因(Thioesterase,PtFATB)可催化水解脂酰基ACP,生成游离脂肪酸和ACP,解除由脂酰基ACP导致的脂肪酸生物合成调节机制中的反馈抑制作用,释放游离脂肪酸[8]。E.coli硫酯酶(Thioesterase I,tesA)是水解长链脂酰辅酶A(C12~C18)形成游离脂肪酸的关键酶[9-10]。本研究在E. coli中分别表达植物来源和内源的硫脂酶基因,将细胞内形成的脂肪酸以游离形式分泌到细胞外,获得产游离脂肪酸的工程菌。
1 材料与方法
1.1 材料与仪器
E. coli K-12、pET30、E. coli BL21(DE3)和拟南芥(Arabidopsis thaliana)组培苗由本实验室提供;Taq DNA polymerase,T4 DNA连接酶购自Takara公司;反转录试剂盒Revert Aid First strand cDNA Synthesis Kit,胶回收试剂盒购自Fermentas公司;PCR引物由上海生工公司合成;游离脂肪酸测定试剂盒购自北京普利莱基因技术有限公司;其他试剂为国产分析纯。MyCycler Thermal Cycler PCR仪、电泳仪和凝胶成像系统为美国BIO-RAD公司生产,Agilent 6897气相色谱由Agilent公司生产。
1.2 实验方法
1.2.1 培养基及培养条件
将-80 ℃甘油保存的E. coli K-12接种于LB液体培养基中[2],37 ℃振荡培养过夜,用于提取基因组DNA。
1.2.2 硫脂酶PtFATB基因的表达
用总RNA提取试剂盒提取拟南芥总RNA,并经Revert Aid First strand cDNA Synthesis Kit试剂盒反转录成cDNA。以拟南芥cDNA为模板扩增PtFATB基因序列,th-L:CATGCCATGGTTTTGAAGCTTTC GTGTAATGTGAC;th-R:CGCGGATCCTTAACTTG AAGGCTTCTTTCTCCAC。PCR扩增条件:94 ℃预变性 3 min,94 ℃ 30 s,58 ℃退火 30 s,72 ℃延伸1.5 min,35个循环。PCR产物与pET30a(+)分别用NcoI/BamHI酶切,构建表达载体pET-PtFATB,并转化至BL21(DE3)感受态细胞中,获得重组菌MX1。
1.2.3 硫脂酶tesA基因的表达
以E. coli K-12 DNA为模板扩增tesA基因序列,tesA-L:CCGGAATTCATGAACTTCAACAATGTTTTCC,tesA-R:ACGCGTCGACTTATGAGTCATGATTTACTAAAGGC。PCR反应条件:94 ℃预变性3 min,94 ℃变性30 s,62 ℃退火30 s,72 ℃延伸2.5 min,35个循环。反应产物和pET30a(+)分别经NcoI/SalI酶切,构建表达载体pET-tesA,并转化至BL21(DE3)感受态细胞中,获得重组菌MX22。
1.2.4 重组酶的诱导表达
将重组菌MX1、MX2分别接种至含50 mL 50 μg/mL Kan的LB液体培养液中,30 ℃诱导表达8~12 h,超声波破壁后进行SDS-PAGE蛋白电泳检测。将重组菌MX1和MX22分别接种在M9基本培养基中,30 ℃诱导12 h后测定代谢产物。总脂含量用游离脂肪酸试剂盒测定,葡萄糖含量用生物传感仪测定。
2 结果与分析
2.1 硫酯酶基因的扩增和表达
重组质粒pMX1和pMX22分别经PCR鉴定、酶切鉴定和测序鉴定,目的片段成功插入到表达载体的相应位点上,获得阳性重组菌MX1和MX22(图1和图2所示)。
如图3所示,SDS-PAGE电泳表明,两种硫酯酶基因在E. coli中成功表达,均可见清晰的条带,tesA基因表达产物约为28.9 KD,PtFATB基因表达产物约为45 KD,与预期结果一致,表明两种硫酯酶基因已成功在E. coli中表达。
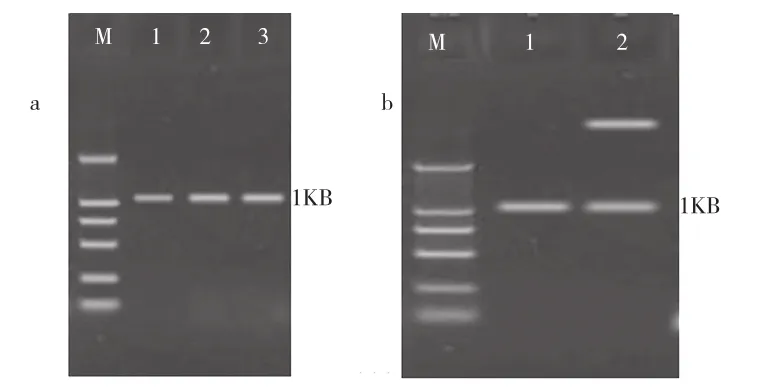
图1 拟南芥硫脂酶基因的PCR扩增和重组质粒构建
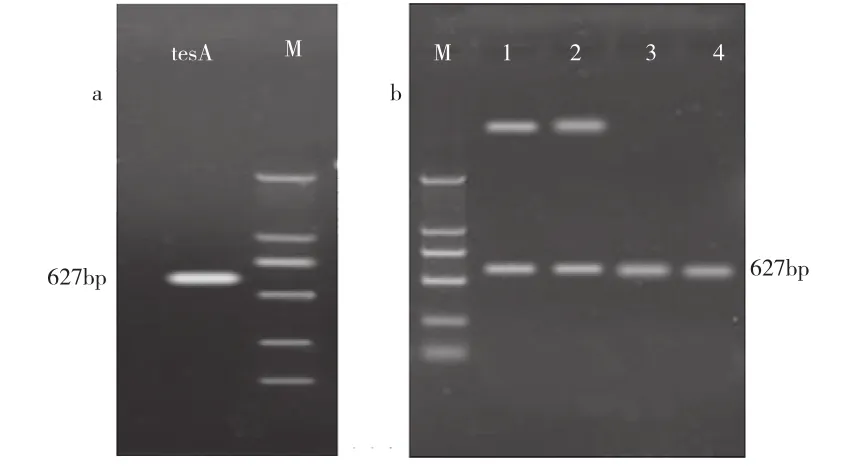
图2 大肠杆菌硫脂酶基因的PCR扩增和重组质粒构建
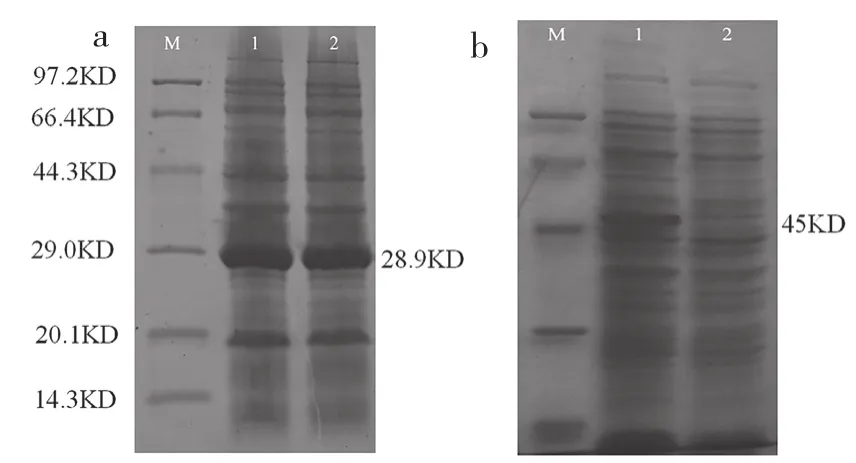
图3 不同来源的硫脂酶基因在大肠杆菌中的表达
2.2 硫脂酶基因功能比较
在E. coli中表达PtFATB和tesA后,获得的胞外游离脂肪酸为33.53 mg/L和25.23 mg/L,为原始菌株的5~6倍,胞外游离脂肪酸与胞内总脂比例为1∶5.8和1∶7.2,游离脂肪酸含量约占总脂的14.71%和12.13%。在天然E. coli中,脂肪酸合成与磷脂合成是紧密相联的过程,细胞内脂肪酸合成的中间产物非常有限,脂酰ACP的大量积累会抑制脂肪酸合成。在硫脂酶水解作用下,可有效解除细胞内的底物抑制现象,E. coli细胞内形成的脂肪酸在硫脂酶水解作用下,释放出游离脂肪酸。两种硫脂酶基因功能验证如表1所示。
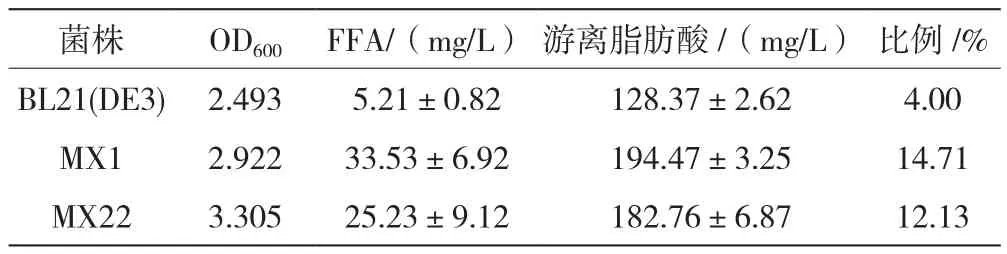
表1 两种硫脂酶基因功能验证
3 结论
在E. coli中分别表达了拟南芥和内源硫脂酶基因,两者均能提高重组菌细胞内不饱和脂肪酸水平,解除产物的反馈抑制作用,脂肪酸以游离形式分泌到外,胞外游离脂肪酸和胞内总脂比例分别为1∶5.8和1∶7,占细胞总脂的14.7%和12.1%。
