两型亚洲小车蝗飞行肌超微结构比较研究
2020-08-25韩海斌高书晶王宁徐林波董瑞文娜仁满都呼
韩海斌 高书晶 王宁 徐林波 董瑞文 娜仁满都呼

摘要 亞洲小车蝗Oedaleus asiaticus B.-Bienko具有长距离迁飞能力,常迁飞为害造成严重损失。掌握其飞行肌结构特征,分析其飞行能力及影响因素,对提高对其监测预警水平具有重要指导作用。本研究采用电子显微镜对群居型和散居型亚洲小车蝗雌成虫背纵肌进行了比较分析。结果表明:群居型和散居型成虫飞行肌具有相似的亚细胞结构,飞行肌的肌原纤维粗细肌丝比例为1∶3,6根细肌丝环绕1根粗肌丝,形成六角形结构。飞行肌的发育和线粒体的形成均是渐进的过程,在不同日龄成虫间存在差异。亚洲小车蝗肌原纤维直径在0.6~1.5 μm,群居型成虫的肌原纤维直径多大于散居型。肌节长度为2.6~4.5 μm,7日龄肌节长度最短,多数日龄群居型成虫肌节长度小于散居型(2.4~3.8 μm)。线粒体在背纵肌内含量占肌纤维线粒体含量的25%~45%,群居型和散居型亚洲小车蝗存在显著差异,群居型7日龄雌成虫线粒体占肌原纤维的比例最高,为52.37%,而散居型的只有32.74%。线粒体含量差异可能是两型亚洲小车蝗成虫飞行能力差异显著的重要原因之一。
关键词 亚洲小车蝗; 群居型; 散居型; 飞行肌; 超微结构
中图分类号: S 433.2
文献标识码: A
DOI: 10.16688/j.zwbh.2019198
Ultrastructure of the flight muscle of female adults in the gregarious
and solitary phases of the grasshopper, Oedaleus asiaticus
(Orthoptera: Acrididae)
HAN Haibin1, GAO Shujing1*, WANG Ning1, XU Linbo1, DONG Ruiwen2, Narenmanduhu2
(1. Grassland Research Institute of Chinese Academy of Agricultural Sciences, Hohhot 010010, China;
2. Siziwang Banner Grassland Station in Wulanchabu League, Inner Mongolia, Wulanchabu 011800, China)
Abstract
Oedaleus asiaticus has the ability to migrate long distance, which has caused negative effects on the areas of destination. Understanding the structure of its flight muscle helps improve the accuracy of monitoring and warning against this pest. The electron microscopy was used to compare and analyze the ultrastructure of flight muscle of female adults in both gregarious and solitary phases of the grasshopper, O.asiaticus. The results showed that the adults of both phases shared similar structures of indirect flight muscle, in which the thick and thin filaments formed a hexagonal lattice with myosin and actin in the ratio of 1∶3. The development of flight muscle and formation of mitochondrion were gradually changeable processes with differences between different day-old adults. The diameters of the myofibril of O.asiaticus were between 0.6 and 1.5 μm. The myofibril diameters of most adults in the gregarious phase were thicker than those in the solitary phase. The length of sarcomere was 2.6-4.5 μm, and the length of sarcomere of 7-day-old adults was the shortest. Most adults in the gregarious phase were shorter than those in the homologous solitary phase (2.4-3.8 μm). The mitochondrion content in myofibril was 25%-45%, which was significantly different between the phases: 52.37%in 7 day-old adults of the gregarious phase and 32.74%in those of the solitary phase. It is inferred that the above differences in mitochondrion content in flight muscle may contribute to different flight abilities in the two phases of O.asiaticus adults: the adults in the gregarious phase can migrate for longer distance.
Key words
Oedaleus asiaticus; gregarious phase; solitary phase; flight muscle; ultrastructure
昆虫的迁飞行为是适应环境而长期进化出的一种生存策略[1],昆虫远距离迁飞寻找食物充足、天敌少等条件适宜环境,增加种群生存繁衍的机会,迁飞害虫常具有突发性、暴发性和毁灭性的为害特征,给农牧业生产造成重大影响[2-3]。适宜的飞行肌结构是昆虫远距离迁飞的基础,昆虫飞行能力的大小与飞行肌结构直接相关[4]。昆虫的飞行肌结构分为两类,同步肌和非同步肌[5],同步肌主要集中在长翅目、蜻蜓目、直翅目、脉翅目和鳞翅目等昆虫上[6]。同步肌在不同种类的昆虫中负担着行走、开掘、弹跳和飞行等多种不同功能,具体功能的不同导致了结构上的分化[7]。肌原纤维是昆虫飞行肌的构成成分,包含多种结构蛋白[8-10],由粗、细两种肌丝沿肌纤维的长轴方向相互穿插、排列,形成规则的空间布局[11-12]。粗、细肌丝通过交叉收缩为昆虫飞行提供强大的动力[13-15]。飞行肌是昆虫飞行的动力基础和结构基础,昆虫不同发育阶段即不同龄期,其飞行肌结构及飞行能力均有很大的差异。飞行肌的发育状况能反映出昆虫的迁飞能力。目前,国内学者已经对小地老虎Agrotis ypsilon、沙漠蝗 Schistocerca gregaria、东亚飞蝗 Locusta migratoria manilensis、意大利蝗Calliptamus italicus等迁飞性害虫的飞行肌超微结构进行了观测[16-19]。
亚洲小车蝗Oedaleus asiaticus B.-Bienko是中国北方典型草原和农牧交错区重要害虫之一[20],多聚集在禾本科草地和过度放牧草原,可以作为内蒙古草原草场退化的指示物种之一[21]。特别是进入21世纪以来,亚洲小车蝗持续暴发成灾,种群数量一般占据整个蝗蟲种群的50%以上,发生严重时种群数量超过90%[22],甚至在内蒙古形成迁飞蝗群,出现大面积起飞和聚集迁移现象,表现与飞蝗相似的习性[23],对农牧业生产造成极大损失。飞蝗在田间有两种存在形态即“群居型”和“散居型”。种群密度低时,蝗虫在田间主要以散居型形态存在,而种群密度较高时,以群居型存在。群居型和散居型蝗虫,共用一套基因,但在体色、行为、飞行能力、生理、形态学上存在明显差异[24]。亚洲小车蝗具有很强的飞行能力,高书晶等研究表明,亚洲小车蝗进行12 h的吊飞测试,群居型最远累计飞行距离可达15 km[25]。在自然生态系统中,亚洲小车蝗存在棕色群居型和绿色散居型,Cease 等研究表明棕色型和绿色型是亚洲小车蝗两型分化的表现型[26]。蝗虫在特定环境下从群居型转变到散居型的形态,这种现象称之为蝗虫的型变。蝗虫的型变被认为是导致其迁飞和暴发成灾的关键因素[27]。本文从超微结构入手研究了亚洲小车蝗背纵肌的组成结构及肌纤维发育动态,明确其飞行肌的飞行能力在不同发育阶段不同,剖析了两型亚洲小车蝗飞行肌结构特征造成飞行能力差异的原理。为全面阐述其迁飞行为机制提供理论依据,有助于完善昆虫迁飞理论和提高测报和防治水平。
1 材料和方法
1.1 供试蝗虫及肌肉取样
试虫亚洲小车蝗采自内蒙古自治区,锡林浩特市镶黄旗和西苏旗,群居型亚洲小车蝗种群采集自小车蝗聚集为害区(≥100头/m2),体色为褐色;散居型亚洲小车蝗种群采集自低密度区(3~5头/m2),体色为绿色,带回实验室饲养至成虫。群居型亚洲小车蝗放入边长为80 cm的正方体养虫笼中饲养,每笼50~60头;散居型放入小笼中单头饲养,用小麦苗及麦麸饲喂。饲养环境条件为温度26~28℃,相对湿度为50%~60%,光照时间为每天14 h。分别取1、4、7、10、13日龄和20日龄的两型亚洲小车蝗雌成虫供试,取中胸背纵肌观测。各日龄群居型和散居型亚洲小车蝗15头作为电镜观察对象。
1.2 样本处理
剥离亚洲小车蝗飞行肌样品,置于2.5%戊二醛溶液中固定,4℃下过夜处理;次日,将样品从固定液中取出,使用漂洗液(pH=7.3的梯度磷酸缓冲液)漂洗,然后用1%的锇酸溶液固定2~3 h;将样品取出用漂洗液漂洗样品3~5次,然后用梯度乙醇溶液对亚洲小车蝗飞行肌样品进行脱水,脱水后使用丙酮清洗掉乙醇溶液,最后使用环氧树脂包埋飞行肌样品并在70℃条件下过夜处理。使用超薄切片机(LEICA EMMZ6)对样品进行切片(厚度约30~50 nm)。经醋酸铀和醋酸铅染色后在电子透射显微镜(Hitachi-7500)下进行超微结构观察。
1.3 显微结构分析
飞行肌肌原纤维的直径为椭圆或多角形横切面的最长轴的长度,纵切面肌节的长度为两Z-带之间的直线距离。横切面的直径和纵切面肌节的长度由显微电镜照片所选取的比例计算得出。根据Steer立体学原理,用已知点数的方格纸覆盖于电子显微照片上,通过计算某细胞器所占的点数与总点数的比来估算出该细胞器在肌纤维中所占的体积分量的方法对亚洲小车蝗飞行肌各结构的体积含量进行估算,然后应用DPS数据处理软件选取Duncan氏新复极差法对各种处理之间的差异进行显著性测验分析。
2 结果与分析
2.1 亚洲小车蝗飞行肌的发育动态
亚洲小车蝗为同步飞行肌,肌原纤维的横切面为椭圆形或者是多角形的结构,肌原纤维被线粒体包围,两者被肌质网分割(图1)。肌原纤维与线粒体的体积比约为1.1∶1.0,大量的线粒体表明,飞行肌对能量的需求较大。亚洲小车蝗成虫羽化后,其飞行肌的肌原纤维、肌质网、线粒体、细胞核、微气管等亚细胞结构均已具备。亚洲小车蝗成虫背纵肌的发育有一个渐进过程,一般成虫羽化后4~5日龄时发育完全。但是飞行肌在成虫阶段也会经历较大的变化。有的变化是定性的,例如肌丝的排列次序,有的变化则是随着成虫发育其长度和宽度等不断变化。这里主要阐述肌原纤维、肌节长度和线粒体的变化。
2.1.1 肌原纤维
2.1.1.1 肌原纤维直径
亚洲小车蝗肌原纤维直径为0.6~1.5 μm,粗肌丝(肌球蛋白丝)的数量为200~700根。6根细肌丝环绕1根粗肌丝,形成六角形结构,每1根细肌丝位于2根粗肌丝的1/2处(图1 b),肌丝排列的精确性,是最显著的特征之一。
不同日龄亚洲小车蝗成虫的肌原纤维直径变化较大。初羽化成虫的肌原纤维直径最小,从1日龄开始至13日龄肌原纤维直径一直处于增大状态,到20日龄肌原纤维直径有所下降(图2)。除10日龄和13日龄之间肌原纤维直径无显著差异外,其余各日龄之间直径存在显著差异(P<0.01);肌原纤维的直径变幅为0.8~1.2 μm。粗肌丝的数目也存在一定差异,但差異不显著(P>0.05)。
2.1.1.2 肌节长度
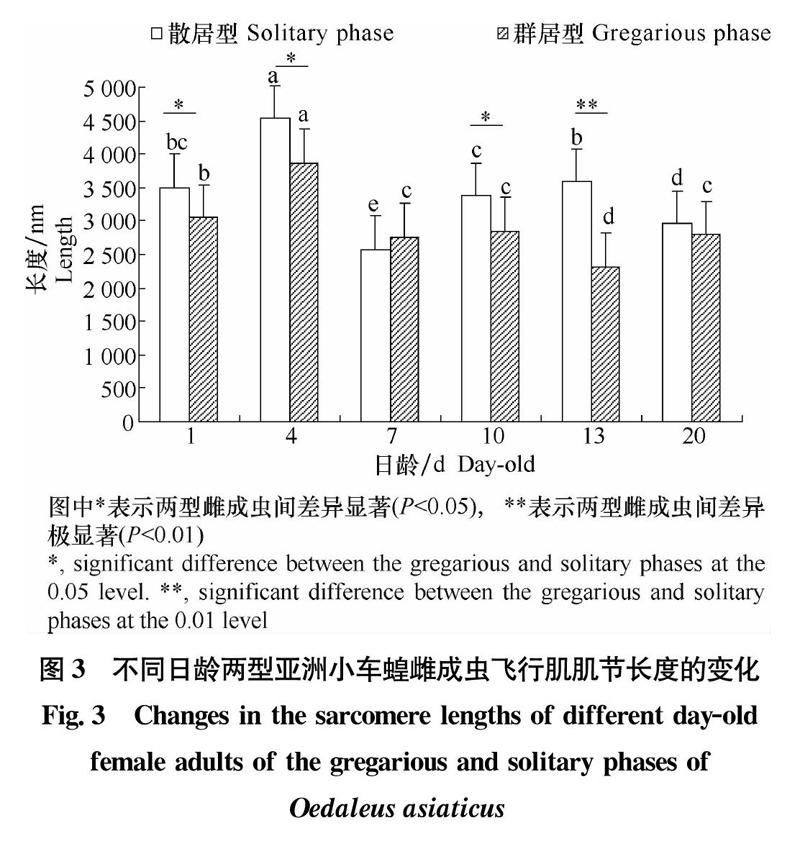
亚洲小车蝗肌原纤维的纵切面为圆柱形(图1),成虫肌节长约2.6~4.5 μm。肌节长度在不同日龄亚洲小车蝗成虫飞行肌的发育过程也存在较大的差异。初期羽化成虫的肌节较短,随着成虫日龄的增加,肌节长度也在不断地变化,在成虫4日龄时肌节长度达到最大(图3)。
2.1.2 线粒体体积分数
亚洲小车蝗成虫羽化初期,背纵肌内的线粒体已经基本成型,形状多为圆形或椭圆形,少数为不规则的形状,边界清楚。随成虫日龄的不同,肌节内线粒体的大小、形状、数量及体积含量等都会发生变化。个体间差异显著。线粒体约占肌纤维总体积的25%~45%。
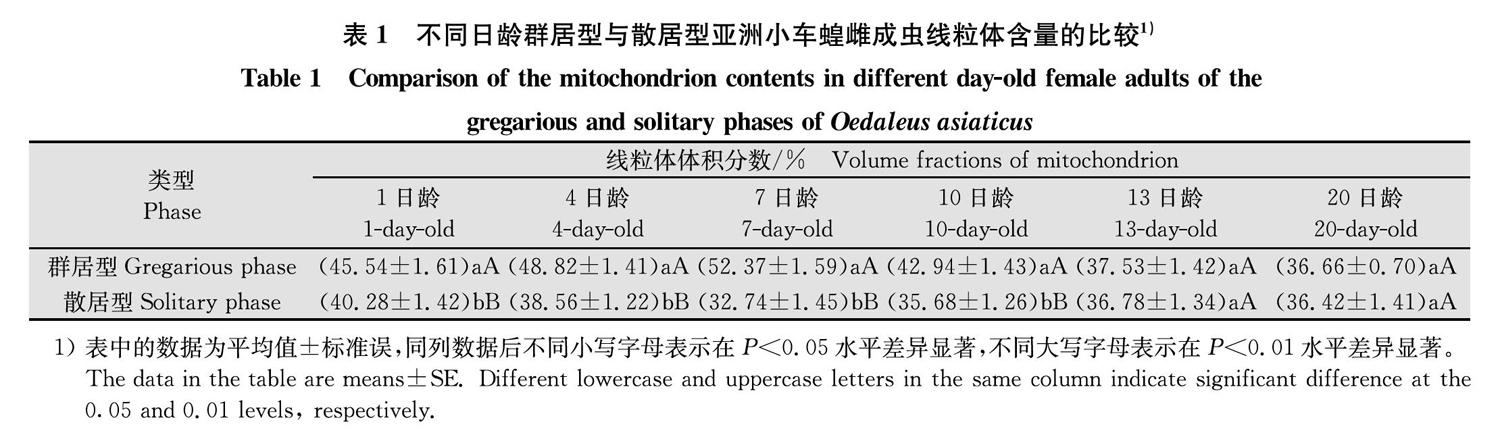
不同日龄成虫背纵肌内线粒体的体积分数随飞行肌发育程度变化见表1。群居型亚洲小车蝗飞行肌中线粒体体积分数随日龄的增加呈先上升后下降的趋势,1日龄成虫线粒体的含量较少,为45.54%;到7日龄时线粒体含量达到最大值,为52.37%,此时肌原纤维间充满线粒体,线粒体内膜结构较紧密;20日龄线粒体含量降到最低,为36.66%,部分线粒体出现破裂的迹象;散居型亚洲小车蝗成虫线粒体体积分数随日龄的增加呈先下降后上升的趋势,7日龄达到最低,为32.74%,10日龄后又有所上升,观察到线粒体变得疏松,出现空洞,少数线粒体膜破裂。
2.1.3 飞行肌其他结构
除了肌原纤维和线粒体外,亚洲小车蝗飞行肌主要结构还有微气管、肌质网和细胞核等。飞行肌细胞内的微气管有两种类型:一类呈椭圆形和长棒槌形,位于肌膜的内侧,微气管内缘呈锯齿状结构。大小为(0.3×1)μm~(2.0×0.8)μm,位置在肌细胞交界处,体积较大(图1)。另一类微气管存在于肌原纤维间(图1),横向分布在线粒体之间,在肌原纤维与线粒体间多呈纵向分布,直径约在0.16 μm。肌质网是由质膜的内膜扩展而成,成虫肌质网不发达,约占肌纤维体积的2.5%。亚洲小车蝗细胞核主要分布在肌膜的内侧(图1),呈椭圆形,不同日龄成虫细胞核的大小不同,平均大小为(5.5×1.1)μm~(8.9×1.3)μm。细胞核在成虫初羽化时较小,发育中期时较大。
2.2 群居型与散居型飞行肌结构的主要差异
2.2.1 肌原纤维差异
群居型和散居型成虫不同发育阶段肌原纤维均有差异,1、10、13、20日龄群居型成虫的肌原纤维直径大于散居型,两者差异显著(P<0.05)。4、7日龄散居型成虫肌原纤维直径略大一些,两者间的差异不显著(P>0.05)(图2)。两型亚洲小车蝗成虫肌原纤维肌节长度在不同日龄之间存在差异(图3)。除7日龄时群居型肌节长度大于散居型外(P>0.05),1、4、10、13日龄群居型的肌节长度小于散居型,且差异显著(P<0.05);13日龄时群居型和散居型肌节长度差异极显著。
2.2.2 线粒体含量差异
群居型和散居型成虫间的线粒体含量在不同发育阶段存在明显差异(表1),其中1、4、7、10日龄散居型亚洲小车蝗雌成虫飞行肌的线粒体含量显著低于群居型雌成虫(P<0.05)。两型之间7日龄时差异达到最大,测定结果显示,群居型线粒体占肌原纤维的比例最高为52.37%,而散居型的只有32.74%。
3 结论与讨论
蝗虫散居型向群居型形态转变是导致远距离迁飞和蝗灾暴发的主要原因,远距离迁飞能力的强弱与昆虫飞行肌的结构特点及发育状况存在直接的关系,飞行肌作为昆虫肌肉系统的重要组成为昆虫远距离迁飞提供强大的基础支撑[28]。肌原纤维和线粒体是亚洲小车蝗飞行肌的重要组成部分,成虫肌原纤维的结构参数有肌原纤维直径、肌节的长度、线粒体在肌纤维中的含量,这些参数很大程度上决定了飞行肌的强度和韧性及飞行时的能源供给[29]。而肌丝排列的有序性、粗肌丝与细肌丝的比例都与肌肉收缩速度有关[7]。电镜检测结果表明,亚洲小车蝗飞行肌的基本结构具有同步飞行肌的基本特征,粗细肌丝相间排列,由6根细肌丝环绕粗肌丝排列成规则的六角形,比例为1∶3,这种结构具有较强的收缩功能和飞行能力。
群居型和散居型亚洲小车蝗飞行肌结构,即纤维直径、肌节长度和线粒体含量经历了从羽化初期到发育盛期的变化过程,从羽化初期到发育盛期,纤维直径增大,肌节长度缩短,线粒体含量增加。10、13日龄是两型亚洲小车蝗飞行肌发育的盛期,后期飞行肌含量下降,发生降解,与不同日龄亚洲小车蝗飞行能力的变化趋势相同[25]。这与对舌蝇 Glossina austeni、飞蝗 L.migratoria 、沙漠蝗S.gregaria飞行肌的研究中得到的结论相同,成虫肌原纤维直径、线粒体体积及飞行肌重量在羽化初期增大,后期飞行肌含量下降,有些发生降解[30-31,17];飞行能力较强的昆虫肌纤维结构特点是肌原纤维直径较粗、肌节长度较短、线粒体较大及含量多,在昆虫远距离飞行中,较粗的肌纤维直径和较短的肌节长度能够增加肌肉收缩频率,提高振翅频率,为昆虫飞行提供良好的动力来源,同时丰富的线粒体可以为飞行提供更多的能量来源[6,18,31]。
比较群居型和散居型亚洲小车蝗飞行肌超微结构的差异可以看出:肌节长度对比,群居型1、4、10、13日龄雌成虫肌节长度短于散居型对应日龄,差异显著;肌原纤维直径对比,群居型1、10、13、20日龄雌成虫肌原纤维直径大于散居型对应日龄,差异显著;线粒体含量对比,群居型1、4、7、10日龄成虫肌原纤维中线粒体含量显著高于散居型,7日龄时两型成虫差异最大,群居型线粒体含量约占肌原纤维52.37%,散居型的仅为32.74%。群居型和散居型亚洲小车蝗飞行肌结构差异主要体现在群居型成虫的肌节长度较短、肌原纤维直径较粗、线粒体体积较大、线粒体含量较高,这些可能是群居型成虫飞行能力显著强于散居型的主要原因之一。刘辉等[32]对东亚飞蝗L.migratoria manilensis Meyen的研究结果显示,群居型具有较强飞行能力,同时,群居型东亚飞蝗的肌节长度较短,肌纤维较粗并含有较多线粒体。发达的肌纤维与丰富的线粒体是小地老虎成虫更适合高速振翅与快速飞行的保障[33]。
昆虫飞行肌在发育过程中是变化的,同时受到环境温湿度、食物丰富度、光照条件、个体拥挤度、风向和风力、防治手段等多种因素的影响[34-35],与其他有迁飞习性的蝗虫相同,亚洲小车蝗具有两型分化,群居型个体较散居型个体有更强的飞行能力,其最大累计飞行距离可达15 km[25],借助气流时扩散距离更远。研究表明,群居型东亚飞蝗最大飞行距离可达35 km[32],群居型意大利蝗借助气流其扩散距离可达120 km[35]。飞行肌是昆虫飞行的动力基础和结构基础,是昆虫特有的肌肉类型。对飞行肌的研究有助于了解迁飞的调控、能量利用等科学问题,飞行肌的发育过程是昆虫生长发育中不可缺少的生理过程,关于迁飞的研究更是昆虫学研究的热点和切入点。国内外科研工作者对飞行肌進行了大量研究,尤其是昆虫飞行肌动力学机制在国外研究较为深入,从物理化学的角度来探究化学能与动能的相互转化,从微观的层面找出昆虫飞行能力可见差异的原因。对于昆虫飞行肌代谢机制的研究也是重要的研究方向[36]。对于亚洲小车蝗飞行肌的研究还需深入,以期能更透彻地剖析其飞行肌的结构、各种蛋白质与迁飞行为的关系等,为合理治理亚洲小车蝗,降低蝗虫迁飞可能性及飞蝗的监测预警提供科学依据。
参考文献
[1] DINGLE H, DRAKE V A. What is migration? [J]. Bioscience, 2007, 57(2):113-121.
[2] RANKIN M A, BURCHSTED J C A. The cost of migration in insects [J]. Annual Reviews of Entomology, 1992, 37:533-555.
[3] JOHNSON C G. Migration and Dispersal of insect by flight [M]. London, UK: Methen, 1969:763.
[4] 丁吉同, 阿地力·沙塔尔, 主海峰, 等. 枣实蝇成虫飞行能力的测定[J]. 昆虫学报, 2014, 57(11):1315-1320.
[5] 杨璞,余海忠,程家安,等. 昆虫飞行肌蛋白质[J].昆虫知识,2005,42(6):726-731.
[6] 罗礼智,李光博.粘虫蛾飞行肌的发育:超微结构特征分析[J].昆虫学报,1996, 39(4):366-374.
[7] PRINGLE J W S. The evolution of fibrillar muscles in insects [J]. Journal of Experimental Biology, 1981, 94: 1-14.
[8] CRAIG R, WOODHEADJ L. Structure and function of myosin filaments [J]. Current Opinion in Structural Biology,2006,16(2): 204-212.
[9] JOHN L C, LORI R N, DAVID W M, et al. Flightin is necessary for length determination, structural integrity, and large bending stiffness of insect flight muscle thick filaments [J]. Journal of Molecular Biology,2010,395(2):340-348.
[10]LING Shanfeng, ZHANG Runjie. Effect of fipronil on brain and muscle ultrastructure of Nilaparvata lugens (Stl) (Homoptera:Delphacidae) [J]. Ecotoxocology & Environmental Safety,2011,74(5):1348-1354.
[11]李红梅. 麦蚜迁飞的生理生化特性研究[D]. 北京:中国农业科学院, 2002:2-18.
[12]韩兰芝,翟保平,张孝羲,等. 甜菜夜蛾飞行肌中与能量代谢有关酶活性[J]. 生态学报,2005,25(5):1101-1106.
[13]MARTYN D A, CHASE P B, REGNIER M, et al. A simple model with myofilament compliance predicts activation-dependent crossbridge kinetics in skinned skeletal fibers [J]. Biophysical Journal,2002,83(6):3425-3434.
