中国北部不同地点樟子松人工林径向生长对气候响应的差异
2020-08-19张劲松何子兵孙守家
雷 帅,张劲松,孟 平,何子兵,程 顺,孙守家 ,*
1 中国林业科学研究院林业研究所/国家林业局林木培育重点实验室, 北京 100091 2 南京林业大学南方现代林业协同创新中心, 南京 210037 3 内蒙古毛乌素沙地开发整治研究中心, 乌审旗 017300 4 河北省塞罕坝机械林场, 承德 068466
根据政府间气候变化专门委员会(IPCC)第五次气候变化评估报告[1],化石燃料的燃烧导致大气CO2浓度从280 μmol/mol增加到400 μmol/mol,全球地表平均气温从1880—2012年升高了0.85℃,并将随着温室气体的持续排放而继续升高,这将导致大部分地区极端高温事件持续时间更长、发生频率更高,而极端低温事件则相对减少[2],森林会遭遇越来越严重的干旱。环境因子的差异对植物径向生长影响显著,其影响程度取决于气候多变和极端事件的发生频率。在干旱和湿润地区,树木生长的限制因子不同,干旱地区树木生长对气候因子的响应强于湿润地区[3]。干旱地区树木生长主要受当年夏季降水的影响[4],夏季降雨比例的增加在一定程度上可以弥补树木由于干旱胁迫等不利因素导致的总增长率的下降[5],但在中欧较湿润地区上年秋季和当年夏季较低的降水对树木径向生长有显著促进作用[6],在高纬度、高海拔的寒冷地区,温度是其主要限制因子[7]。
树木年轮包含了树木历年的气候变化信息,成为追溯树木对过去气候长期变化响应的重要材料[8-9],受到广泛的关注[10-11]。樟子松天然分布于我国呼伦贝尔沙地和大兴安岭北部,在“三北”防护林工程中被大规模引种栽植,具有较强的生态适应性,面积已超过3.0×105hm2。作为“三北”地区主要造林树种之一,在经过初期良好的生长后,开始出现大面积衰退现象[12-13],因此,对樟子松在不同地区适应性研究非常重要。先前对樟子松的研究多集中在同一地点树木径向生长对气候的响应[14],例如红花尔基、海拉尔等地樟子松生长主要受降雨影响[15-16],大兴安岭、漠河、永安山等地的树木生长主要受温度驱动[17-19],同一地区樟子松生长也会受到小气候差异的影响[20],但对不同地点相同密度樟子松对气候的响应关系认识不足,制约了对气候差异下樟子松人工林生长、衰退和更新状况的预测。
本研究地点在河北省塞罕坝机械林场和内蒙古鄂尔多斯市毛乌素沙地研究中心,二者环境不同且有相同密度的樟子松林,可消除林内竞争的影响。不同地点相同密度樟子松轮宽及胸高断面积增量(Basal area increment,BAI)的变化是否存在差异?樟子松早材和晚材所占比例变化趋势是否一致?影响樟子松径向生长的主导气象因子是否相同?弄清这些问题对于探求樟子松人工林对气候响应有重要意义。因此,本研究利用树木年代学方法对不同地点相同密度的樟子松人工林进行测定,分析其径向生长的差异以及对气候因子的响应,旨在明确:(1)樟子松轮宽及BAI的差异性;(2)早材晚材所占比例的变化趋势;(3)影响樟子松径向生长的主导气候因子。通过分析樟子松径向生长差异和导致差异的原因,加深对不同地点樟子松适应性的理解,为制定适当的樟子松人工林经营管理理论提供依据。
1 研究方法
1.1 研究区概况
图1为研究区概况图,毛乌素沙地位于鄂尔多斯高原向陕北黄土高原的过渡地区,属于荒漠草原, 取样点位于内蒙古鄂尔多斯市乌审旗境内毛乌素沙地整治开发研究中心(38°57′—39°61′N,109°21′—109°17′E,海拔1283 m,以下简称毛乌素),多年平均气温6.8℃,降水量350—400 mm,蒸发量2200—2800 mm,平均风速3.4 m/s,无霜期113—156 d,温带大陆性季风气候。土壤PH为8.44,总碳含量为6.91 g/kg,总钾含量为0.68 g/kg,总磷含量为0.45 g/kg,总氮含量为0.31 g/kg。

图1 实验样地分布图[13] Fig.1 Experimental plot distribution
塞罕坝位于河北省承德市围场满族蒙古族自治县境内,分为坝上、坝下两部分,坝上是内蒙古高原南缘。取样点位于坝上塞罕坝机械林场(41°35′—42°40′N,116°32′—118°14′E,海拔1553 m,以下简称塞罕坝),多年年均气温-1.3℃,极端高温33.4℃,极端低温-43.3℃,积雪最长达7个月,降水量460.3 mm,无霜期64 d,是典型的半干旱半湿润寒温性大陆季风气候。土壤PH为6.29,总碳含量为3.02 g/kg,总钾含量为0.41 g/kg,总磷含量为0.2 g/kg,总氮含量为0.32 g/kg。
1.2 样本采集
在2个取样点分别随机设置20 m×20 m的樟子松人工林样地,测定密度、树高、胸径等参数(表1),在树干胸径(1.3 m)处,使用生长锥钻取树芯,在每株东-西、南-北方向各取1根,采集完毕后,使用玻璃胶及树木创伤修复液将树木创口封住,一个采样点取40棵树共80根样芯,树芯采集后将样品及时放入专用取样盒中带回实验室,放置于通风条件下自然晾干。

表1 样地基本情况统计表
1.3 树轮宽度测定及年表建立
先使用低目数砂纸将样品适度打磨,再用高目数的砂纸抛光,使用Lintab 6年轮分析仪(Rinntech 公司,德国)和TASP-Win软件测定早晚材年轮宽度,全材轮宽由早晚材求和所得,精确至0.01 mm,通过折线图法进行交叉定年,并使用COFECHA程序检验,以确保定年和测量准确。由于样本树龄较短,研究主要集中在树木生长的高频变化上,故应去除树木随年龄增长的生长趋势和其他非一致性扰动导致的非气候信号,而突出其中的气候信号[21],利用ARSTAN程序建立年表,去趋势方法为负指数函数,该方法可减少树间竞争、地形、地貌等因素对年表指数拟合的影响,最终得到标准化年表(Standard chronology,STD)、自回归年表(Arstan chronology,ARS)和差值年表(Residual chronology,RES)。本研究选用RES进行树木生长与气候间的相关分析,RES是在STD的基础上,考虑到森林内部树与树的竞争以及可能存在的人类活动导致的树轮宽度序列的低频变化,以及时间序列的自回归模式对STD进行拟合并再次标准化,去掉树木个体特有的前期生理条件对后期生长造成的连续性影响而建立的一种年表,它只含有群体共有的高频变化。
1.4 胸高断面积增量
当树轮宽度相对稳定或变窄时,树木生长量可能继续增大[22],为了保留气候条件对树木生长影响的信息,本文使用BAI来评估研究区内樟子松人工林对气候变化的响应,公式如下[23-24]:
式中,R代表树木半径,n表示年轮形成年份。
1.5 气候数据与干旱指数
气象数据来源于离采样点较近的国家标准气象台站(毛乌素38°16′N,109°47′E,海拔1157 m;塞罕坝42°18′N,117°25′E,海拔1546 m),各选取2站点樟子松生长期间的气温、降水、相对湿度等数据。标准化降水蒸散发指数(Standardized precipitation evapotranspiration index,SPEI)采用三参数的Log-Logistic 概率密度函数f(x)来描述月降水与月潜在蒸散量差值的变化,得到月水分平衡量的概率分布函数F(x),经正态标准化处理后获得[25]。根据降水距平百分率、SPEI 和标准差d指数将干旱划分为5个等级(表2)。

表2 干旱指数等级划分
1.6 数据分析
樟子松树轮生长除受当年气象因子的影响外,还受到上一生长季诸多气候因子的影响,因此进行树轮气候响应分析时的时间跨度为前一年6月份到当年10月份。使用R软件包“Treeclim”分析RES与气候因子的关系[26],Origin 2017绘图,AMOS 20(IBM,美国)建立结构方程模型,结构方程模型结果图中,气候因子如平均降雨、平均温度等设为原因变量,全材(Ringwood,RW)、早材(Earlywood,EW)和晚材(Latewood,LW)设为结果变量,在标准化结果中,原因变量到结果变量路径线上的系数代表作用效应的大小。
2 结果与分析
2.1 气候因子变化
图2为研究区气候变化趋势图,毛乌素多年年均温8.25—10.57℃,年均降水量约410 mm,年均相对湿度40.12%—62.73%。年SPEI范围-1.82—2.05,波动较大,毛乌素大体属于中—重度干旱,SPEI每10年上升0.46,干旱情况在逐渐缓解。从月尺度上看,降水主要集中在7—8月,最高温度出现在7月,相对湿度4月份最低,8月份最高,7月份 SPEI最低。塞罕坝多年平均温-2.95—0.89℃,年均降水量约465 mm,年均相对湿度56.78%—66.38%,年SPEI范围-1.73—2.14,波动较大,总体上塞罕坝大体属于中—轻度干旱,SPEI每十年下降0.004。从月尺度上看,降水主要集中在7—8月,最高温度出现在7月,相对湿度4月份最低,8月份最高,月SPEI波动较小,达到无旱水平。

图2 毛乌素和塞罕坝平均气候趋势图Fig.2 Average climate trend chart of Mu Us and Saihanba sites
毛乌素和塞罕坝多年气象数据表明二者存在明显差别,毛乌素降水呈明显上升趋势,每10年上升约8.61 mm,尤其是最近10年,增加较为明显,塞罕坝降水基本保持稳定。毛乌素温度呈明显升高趋势,每10年约上升0.40℃,塞罕坝温度呈不显著上升趋势,每10年约上升0.20℃。毛乌素多年均温高于塞罕坝约8℃左右。毛乌素空气相对湿度呈明显下降趋势,每10年约下降3.40%,而塞罕坝缓慢升高,每10年约上升0.50%。毛乌素SPEI呈明显线性升高趋势,干旱情况在逐渐缓解,塞罕坝则相对稳定。
2.2 樟子松轮宽变化
图3为樟子松年际轮宽变化趋势图,结果显示,2个试验点的轮宽变化趋势明显不同,毛乌素樟子松轮宽呈“升-降”趋势,而塞罕坝呈线性下降趋势。毛乌素樟子松RW、EW和LW的轮宽变化趋势基本一致,从近30年的轮宽拟合曲线来看,樟子松RW、EW和LW轮宽在1989—2000年期间上升,轮宽从1989年的2.34、1.51、0.83 mm分别上升到2000年的3.02、1.98、1.04 mm,分别增加29.06%、31.13%、25.31%,2000—2018年期间下降,2018年下降到1.52、1.27、0.25mm,分别下降49.67%、35.86%、75.96%。结合气象及轮宽实测数据分析,毛乌素2000年属于重度干旱,RW轮宽比1999年下降了6.49%,LW轮宽下降了23.36%,而EW轮宽上升16.45%,表明樟子松晚材径向生长受当年环境因素影响较大。早晚材轮宽百分比图结果显示,毛乌素樟子松早晚材轮宽波动相对较小,EW/RW比值范围为59.65%—71.36%,均值为64.26%,呈微弱下降趋势,30年约下降0.64%,EW/RW比值范围为29.38%—41.64%,均值为35.74%,呈微弱上升趋势。

图3 毛乌素和塞罕坝樟子松轮宽变化Fig.3 Ring-width variation of Pinus sylvestris var. mongolica of Mu Us and Saihanba sitesRW: Ringwood; EW: Earlywood; LW: Latewood
塞罕坝樟子松RW、EW和LW轮宽呈相似的变化趋势,近40年的轮宽拟合曲线显示RW、EW和LW均呈显著下降趋势(P<0.05),轮宽分别从1979年的3.99、2.73 、1.26 mm下降到2018年的1.25、0.81、0.44 mm,分别下降了68.67%、70.33%、65.08%。塞罕坝在2000年和2006年属于重度干旱,RW轮宽比前一年分别下降了6.60%和1.63%,LW轮宽比前一年分别下降了46.30%和27.27%,而EW轮宽分别上升9.13%和12.41%。早晚材轮宽波动相对较大,EW/RW值范围为55.35%—79.67%,均值为69.32%,呈明显下降趋势,40年约下降9.60%,LW/RW值为21.32%—45.78%,均值为30.68%,呈明显上升趋势。
2.3 樟子松BAI变化
图4为研究区樟子松BAI年际变化趋势图,结果显示,毛乌素樟子松近30年生长期间RW、EW和LW的BAI有相似的变化趋势,但年际间BAI波动较大。拟合曲线值显示RW、EW和LW的BAI从1989年的1.14、0.71 cm2和0.43 cm2升高到2011年的10.95、6.28 cm2和4.67 cm2,分别上升860.53%、784.51%、986.05%,2011—2018年期间下降,到2018年下降到9.14、6.16 cm2和2.98cm2,分别比2011年下降16.53%、10.19%、36.19%。毛乌素在2000年属于重度干旱,BAI实测值分析,RW比1999年下降了15.28%,LW下降了42.42%,而EW上升了2.17%。早晚材BAI比值图结果显示,毛乌素樟子松早晚材BAI波动相对较大,EW/RW波动范围为43.36%—67.45%,均值为58.65%,呈下降趋势,30年约下降3.60%,LW/RW波动范围为26.31%—57.43%,均值为41.35%,呈上升趋势。

图4 毛乌素和塞罕坝樟子松BAI变化Fig.4 BAI variation of Pinus sylvestris var. mongolica of Mu Us and Saihanda sites
塞罕坝樟子松RW、EW和LW的BAI均呈显著的二次曲线变化。拟合曲线值显示RW、EW和LW的BAI从1979年的0.51、0.28 cm2和0.23 cm2升高到1998年的8.98、5.34 cm2和3.64 cm2,分别增加了1660.78%、1807.14%、1482.61%。在1998—2018年开始下降,到2018年为5.86、3.93、1.93 cm2,分别下降34.74%、26.41%、46.98%。塞罕坝在2000和2006年属于重度干旱,BAI实测值分析,RW比前一年分别下降了18.38%和8.88%,LW比前一年分别下降了73.03%和46.64%,而EW比前一年分别上升了3.48%和12.56%,这与干旱胁迫下的轮宽变化相似,说明干旱胁迫对晚材的形成与生长影响较大。早晚材BAI比值图结果显示,塞罕坝樟子松早晚材BAI百分比波动相对较大,EW/RW波动范围为55.36%—74.79%,均值为68.32%,呈明显下降趋势,40年约下降9.60%,LW/RW波动范围为22.76%—45.64%,均值为31.68%,呈明显上升趋势。
2.4 年表特征统计
研究结果的准确与否取决于年表质量的高低,平均敏感度(Mean Sensitivity,MS)、样芯间相关系数(Mean inter-series correlation,RBAR)、信噪比(Signal to noise ratio, SNR)、样本总体代表性(Expressed population signal,EPS)等均用来评价年表质量。RBAR是指每个采样点内测得的各样芯年轮宽度序列间的相关性,相关性越高代表不同样芯间的年轮宽度变化具有较好的一致性,数据质量越好[27]。MS和SNR反映相邻年轮之间年轮宽度的变化情况,主要代表气候的短期变化或高频变化,MS大于0.15 代表年表质量较高[28-29]。EPS是指所采集的样本对整个区域的代表程度,通常EPS大于0.85 的年表质量较高[30]。
表3年表统计特征值显示,年表RBAR都在0.5以上,表明同一采样点内树木径向生长具有共同的气候限制因子,导致年轮宽窄的同步性变化。在本研究的所有年表中,平均敏感度基本达到了0.15,SNR比较高,其值越大,表示树木年轮中包含的气候信号越多,说明样本所包含的气候信息量较大,基本符合与气象要素的相关分析要求。所有年表的EPS均大于0.9,表明所采样本对区域人工林变化具有较好的代表性。

表3 年表统计特征值
从图(5)RES趋势图中可以看出,全轮与早材RES年表在高频变化上更为一致,说明全轮和早材年表中所包含的气候信息可能更为相似。各采样点年表在某些年份表现出生长变化的一致性,如毛乌素的RW、EW、LW在1999、2005和2016年均出现峰值,在2010、2012和2006年均出现谷值;塞罕坝的RW、EW、LW在1991和2011年均出现峰值,在1983、1986和2002年均出现谷值。RBAR系数的大小表明,早、晩材序列的变化均具有较好的一致性,但是略低于全轮(表3)。三者比较,全材各序列间变化的同步性最好,早、晚材年表的MS、EPS均比较高,表明人工林樟子松早、晚材对气候变化敏感,包含丰富的气候信息,早晚材宽度变化适用于树木年轮气候学分析。
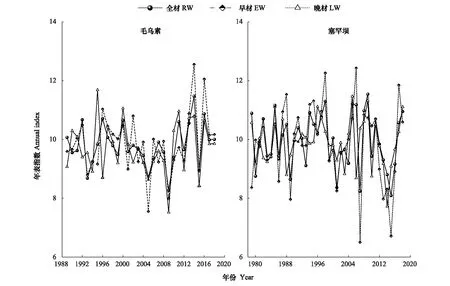
图5 毛乌素和塞罕坝年表指数趋势图Fig.5 Chronological Index trend of Mu Us and Saihanba sites
2.5 RES与月气候因子的关系

图6 差值年表与气候因素相关系数图Fig.6 Correlation coefficients between the index of the residual chronology(RES)and climatic factorsPRE:平均降雨 Average precipitation;T:平均温度 Average temperature; SPEI:干旱指数 Standardized precipitation evapotranspiration index;RH: 相对湿度 Relative humidity; Tmin: 最低温度 Minimum temperature; Tmax: 最高温度 Maximum temperature; RW:全材;EW:早材; LW: 晚材
图(6)表示差值年表与气候因素相关系数图,结果表明,毛乌素樟子松RW的RES与上年7月及当年4、7月份的平均降雨显著正相关,与上年8月及当年3、8月份平均温度显著正相关,与当年7月份相对湿度显著正相关,与上年及当年7月份SPEI显著正相关,与上年12月和当年3月份最低温度显著正相关,与上年11月及当年6月份最高温度显著负相关。EW的RES与上年7月及当年4、7月份的平均降雨显著正相关,与上年8月及当年3、4、8、9月份平均温度显著正相关,与上年8月份相对湿度显著负相关,与上年及当年7月份SPEI显著正相关,与上年12月和当年3、4月份最低温度显著正相关,与上年6月及当年6月份最高温度显著负相关。LW的RES与上年及当年7月份的平均降雨显著正相关,与上年8月及当年3、8月份平均温度显著正相关,与当年7月份SPEI显著正相关,与当年3月份最低温度显著正相关,与当年8月份最高温度显著负相关。
塞罕坝樟子松RW的RES与上年及当年7、8月份平均降雨显著正相关,与上年9月相对湿度显著负相关,与上年7、8月SPEI显著正相关;与当年8月份最低温度显著正相关,与5月份最高温度显著负相关。EW的RES与上年7月及当年5、8月份平均降雨显著正相关,与上年8月份相对湿度显著正相关,与上年7、8月及当年5、8月SPEI显著正相关,与上年7月及当年5、7、8月份最低温度显著正相关,与上年10月和当年3、5月份最高温度显著负相关。LW的RES与上年及当年7、8月平均降雨显著正相关,与上年7月及当年3、7月SPEI显著正相关,与上年及当年7月份最低温度显著正相关,与当年5月份最高温度显著负相关。
2.6 RES与年气象因子的关联
图7表示毛乌素和塞罕坝樟子松RES与气象因子的路径分析,利用AMOS软件对毛乌素与塞罕坝樟子松构建了结构方程模型[31]。由于两个实验地点影响樟子松的气象因素较多,在一个结构方程模型图中对RW、EW、LW的影响线路繁杂,故图7表示的是简化了的结构方程模型图,图中系数表示气象因素对樟子松总效应的大小。结果显示,在年尺度上SPEI对毛乌素樟子松RW有极显著负影响(P<0.01),效应达到0.92,平均温度对毛乌素樟子松RW有极显著负影响(P<0.01),效应达到0.77,平均降雨对RW有显著正影响(P<0.05),效应达到0.31,平均降雨对EW有极显著正影响(P<0.01),效应达到0.44,SPEI对EW有极显著负影响(P<0.01),效应达到0.99,最低温度对LW有极显著正影响(P<0.01),效应达到0.49,平均温度对LW有极显著负影响(P<0.01),效应达到0.84。塞罕坝SPEI对RW有极显著负影响(P<0.01),效应达到0.58,平均温度对RW有极显著负影响(P<0.01),效应达到0.66,平均降雨对EW有极显著正影响(P<0.01),效应达到了0.72,SPEI对EW有极显著负影响(P<0.01),效应达到0.58,平均温度对LW有极显著负影响(P<0.01),效应达到了0.61,最低温度对LW有极显著正影响(P<0.01),效应达到了0.57,平均降雨对LW有极显著负影响(P<0.01),效应达到了0.75。
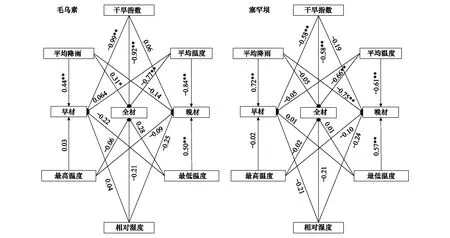
图7 毛乌素和塞罕坝樟子松差值年表与气象因子的路径分析Fig.7 Path analysis diagram RES of Mongolian pine trees and meteorological factors of Mu Us and Saihanba sites
3 讨论
3.1 轮宽及BAI差异分析
气象因子的差异对树木径向生长影响显著[32-33],高温和干旱对植物生长产生不同程度的影响[34-36]。研究区毛乌素和塞罕坝具有明显气候差异性,通过对比两个试验点樟子松径向生长,对不同气候环境下樟子松的适应性研究有重要意义。对比发现,毛乌素樟子松轮宽呈波动的二次曲线变化,塞罕坝则呈线性下降,说明毛乌素多变的气候环境对樟子松的交互影响更为显著,其樟子松在生长过程中对温度和水分的敏感性更高;而塞罕坝的樟子松随着树龄的增大,樟子松的径向生长速率在逐步减缓,因此轮宽在逐年变窄,独特的低温高湿环境使樟子松的生长波动较小。两地干旱胁迫均使樟子松轮宽及BAI显著降低,说明干旱胁迫会严重抑制樟子松的径向生长。从两地樟子松逐年BAI增长量分析,由于两地樟子松的树龄均较小,只有30—40年,而樟子松天然林中值年龄大约可达230年,最大年龄可达350年左右[37],樟子松人工林也可达50年左右[38],因此,两地樟子松BAI逐年增长量变化趋势并不能真实反映樟子松生长有无衰退迹象。
从早晚材百分比变化分析,两地轮宽和BAI百分比拟合线趋势基本同步,毛乌素早晚材百分比上下波动较大,但拟合线相对平滑,上下浮动的范围相对稳定,说明毛乌素波动的气象因素对早晚材的影响均较大。塞罕坝早晚材百分比波动相对较小,但明显EW百分比向下浮动,LW百分比向上浮动,导致EW百分比显著线性下降,LW百分比显著线性上升,所以塞罕坝稳定的气候条件更利于后期LW的生长。毛乌素干旱胁迫使LW百分比下降,EW百分比升高,说明干旱胁迫对LW的生长产生了抑制作用,从而使RW轮宽及BAI减少,这也反映出当年樟子松EW可能是使用上季同化的碳形成,而LW主要依赖于其当年生长季积累的碳[39-40]。
3.2 气象因子响应差异分析
不同地点的樟子松对月气象因素的响应不同,RES与月气象因子的相关性分析表明,毛乌素樟子松径向生长与4、7月的降雨和3、8月的均温及低温呈正相关关系,与上年11月及当年6月最高温度呈负相关关系,原因可能是上年11月气温过高,会降低降雨和降雪的频率,使土壤水分减少,影响来年樟子松的初期生长,而6月属于生长季前期,高温加快了土壤水分蒸发,提高了蒸汽压差,限制了树木的生理代谢活动[41]。3、4月即初春平均温度的回升及降雨的增多对樟子松的初期形成层细胞的分裂有显著促进作用[42-43],7、8月即生长季初夏到初秋是树木生长最旺盛的时期,也是季风降水的高峰期,最低温度的回升有利于树木的光合作用,从而产生足够的光合产物供给树木生长,而温度过低则限制其光合[44-45]。塞罕坝海拔高温度低,常年处于低温高湿环境,最低温度与最高温度的变化,会显著影响樟子松的光合作用,从而改变其径向生长速率[46]。塞罕坝樟子松径向生长与上年7、8月和当年5、7、8月的平均降雨呈显著正相关,与上年7月及当年8月SPEI呈显著正相关,与当年5月最高温度呈显著负相关。原因可能是樟子松天然分布于寒冷的呼伦贝尔沙地,喜欢冷凉气候[47-48],5月最高温度的升高不利于樟子松内在水分积累,樟子松对塞罕坝的天然低温有明显适应性。5、7、8月份降雨的充沛能有效补给土壤水源,供给植物的快速生长,因此,与降雨及SPEI均呈显著正相关。
树木的径向生长主要受树木本身遗传特性、立地条件和气候条件等外部环境条件的综合作用[49],AMOS结构方程模型表明,两地年SPEI和年平均温度对樟子松RW径向生长影响显著,且均为负效应,毛乌素年平均降雨对RW产生显著正效应,原因可能是年SPEI受年平均温度和年平均降雨的影响,年平均温度与年SPEI对两地樟子松RW均产生了负效应,而年平均降雨却相反,说明影响两地年SPEI较大的是年平均温度,毛乌素樟子松RW生长的限制因素是年平均温度与年平均降雨,塞罕坝RW生长的限制因素是年平均温度,且两地年平均温度的升高均不利于樟子松的径向生长[50-51]。两地年SPEI和年平均降雨对樟子松EW径向生长影响显著,且年SPEI为负效应,年平均降雨为正效应,上述可分析年SPEI的负效应可能由年平均温度引起的干旱胁迫产生,年平均温度对EW的影响并未达到显著水平,因此,EW的限制性因素是年平均降雨,且与其正相关[52],年平均降雨对两地EW产生了极显著正效应,说明两地樟子松EW的生长对水分依赖性较强,EW管胞生长与上一年的冬季降水有关,生长后期的管胞受当年春季中期降水的影响[39]。两地年平均温度和年最低温度对樟子松LW径向生长影响显著,且年平均温度为负效应,年最低温度为正效应,说明樟子松适宜生长于冷凉环境,这与樟子松引种地有关[47-48]。塞罕坝的年平均降雨对LW的生长产生了负效应,原因可能是LW生长发生在8月份之后, LW的形成与生长主要与形成层细胞分裂缓慢和木质部结构致密有关。晚材形成主要利用当季积累的碳,此时塞罕坝的平均温度已经降至8℃以下,而平均降雨的增多并不能补偿光合作用的亏缺,相反,后期温度的进一步降低引发的冻害则加剧了这种状况[53],因此,毛乌素LW的限制性影响因素是年平均温度和年最低温度;塞罕坝LW的限制性影响因素是年平均温度、年最低温度和年平均降雨。
综合分析,我国樟子松人工林耐寒性强,不苛求较高的土壤水分,能适应土壤水分较少的山脊及向阳山坡,以及较干旱的砂地及石砾砂土地区,因此,较适宜种植于冷凉气候环境下的“三北”地区。但是,也有一些限制性条件需要注意,一些雨量较少而生长季温度较高的干旱地区,特别是遇到重度干旱年份,需要进行适当的土壤增湿处理,一些雨量较高但温度很低的高寒地区,冬季要注意樟子松幼苗的冻害防护工作。
4 结论与展望
毛乌素樟子松轮宽呈显著二次曲线变化,塞罕坝樟子松轮宽呈显著线性下降,两地樟子松EW生长对全材贡献较大。两地樟子松径向生长与生长季前期及生长季的月平均降雨和月最低温度呈正相关,晚冬及生长季月最高温度对其负相关,毛乌素樟子松在生长过程中对温度的敏感性比塞罕坝更高。两地樟子松RW生长的共同限制因子是年平均温度和年SPEI,EW的限制性因子是年SPEI和年平均降雨,LW的限制性因子是年平均温度和年最低温度。塞罕坝樟子松径向生长对气象因子响应与毛乌素相似,但略有差别,塞罕坝年平均降雨对LW产生极显著负效应,但对EW未达到显著性水平,且SPEI对塞罕坝樟子松RW和EW产生的干旱胁迫效应明显小于毛乌素。总之,两地年平均温度、年最低温度、年SPEI和年平均降雨是樟子松径向生长的显著限制因子,但各气象因子对其总效应差异较大。
樟子松作为“三北”防护林的重要造林树种之一,如何应对近年来出现的大面积衰退问题,对其不同区域环境下适应性探究势在必行。樟子松人工林衰退不仅与本身消耗水资源有关,也与其对不同区域气候适应性有关,因此,如何解析区域尺度水分亏缺形成机制,探究其生态水文过程,如何确定樟子松人工林应维持的适宜规模和最优分布格局,探究其最适种植、最优生长,如何解析树木应对干旱,特别是极端干旱胁迫的恢复能力,依旧是研究中的难题。本文对不同气候环境下樟子松生长差异及其与气候因子的关联进行了探究,但未对其生理适应性机理进行深入解析,樟子松衰退的原因非常复杂,可能是林内竞争或气候变化导致的,也可能是二者协调作用共同造成的,需要后期研究中加入更多因素综合考量,如养分、水分和竞争要素等,开展气候、竞争和碳水耦合等机制的解析。
