中街山列岛海域鱼卵、仔稚鱼群落结构特征及其与环境因子的关系
2020-08-19樊紫薇蒋日进李鹏飞张琳琳
樊紫薇,蒋日进,*,李 哲,李鹏飞,梁 君,张琳琳,印 瑞
1 浙江海洋大学海洋与渔业研究所,舟山 316021 2 浙江省海洋水产研究所,农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,舟山 316021
鱼卵、仔稚鱼是海洋食物链中的重要环节之一,在海洋生态系统系统,两者是重要的被捕食者,同时仔稚鱼也是捕食者,它们的存活数量是鱼类资源补充和渔业资源持续利用的基础,其数量变化直接影响渔业资源量的变动[1- 2]。同时,为评估鱼类资源量及保持渔业资源的可持续利用提供了基础资料[3],也为鱼类资源养护管理和渔业生态环境的保护提供了参考依据[4- 5]。
中街山列岛海域位于舟山渔场的中心区域,属亚热带季风气候,历史上素有“鱼仓”之美誉。历史上是曼氏无针乌贼重要的产卵场,也是大黄鱼进入岱衢洋、大戢洋产卵场的重要通道[6],渔业资源丰富,但近年来由于气候变暖、沿岸设施工程的增加及捕捞压力过大等综合原因,导致渔业资源衰退。该海域受浙江沿岸流和台湾暖流的交汇影响,同时长江、钱塘江等大陆径流带来了丰富的营养物质,使之成为众多海洋生物繁殖、摄食和生长的重要栖息地[7]。有关其他海域的鱼卵仔稚鱼群落结构已有相关研究报道,刘鸿等[8]对黄海中部近岸春夏季鱼卵、仔稚鱼群落结构特征进行研究,得出鱼卵丰度和分布与经度呈负相关、与表温呈正相关,但与其他环境因子如纬度、表盐和水深等无显著相关性。仔稚鱼丰度和分布与上述环境因子均未表现出显著相关性;刘守海等[9]对2005年、2008年、2009年和2011年夏季长江口水域鱼卵、仔稚鱼年间变化进行分析,结果表明该水域鱼卵和仔稚鱼2005年夏季种类数最多,2008年夏季种类数最少;杨艳艳等[10]调查发现莱州湾春、夏季鱼卵、仔稚鱼优势种季节变化明显,且盐度对鱼卵数量影响较为明显, 温度对仔稚鱼数量影响较为明显。目前,有关中街山列岛海域的鱼卵、仔稚鱼群落结构及其与环境因子关系的相关研究尚未报道。本文基于2010—2012年中街山列岛海域定量采样数据,分析了该海域鱼卵、仔稚鱼的种类组成、数量空间分布、物种多样性以及与环境因子的关系等方面,以期为该海域渔业资源的科学管理和海洋生态的可持续发展提供科学依据。
1 材料和方法
1.1 调查站位及样品采集
在中街山列岛海域设置10个站位(图1),于2010年5月—2012年2月四季(2月、5月、8月和11月)进行了8个航次的鱼卵、仔稚鱼调查。鱼卵、仔稚鱼采集方法依据《海洋调查规范 第6部分:海洋生物调查》(GB/T 12763.6—2007)[11],采用大型浮游生物网(口径80cm、长280cm、网目尺寸0.505cm)逐站进行表层水平拖网采样,每站拖10min,拖速为2 n mile/h;样品的垂直采集采用大型浮游生物网由底到表垂直拖取,拖速0.5m/s。对温度、盐度、悬浮物等相关环境因子数据采用多功能水质检测仪(JFE-AAQ171)进行同步采集。平拖样品用无水乙醇固定保存,垂拖样品用5%福尔马林溶液固定,带回实验室,在显微镜下根据鱼卵、仔稚鱼的形态特征进行种类鉴定,并计数。对于不确定的种类采用分子测序的方法,尽可能鉴定到最低分类等级。
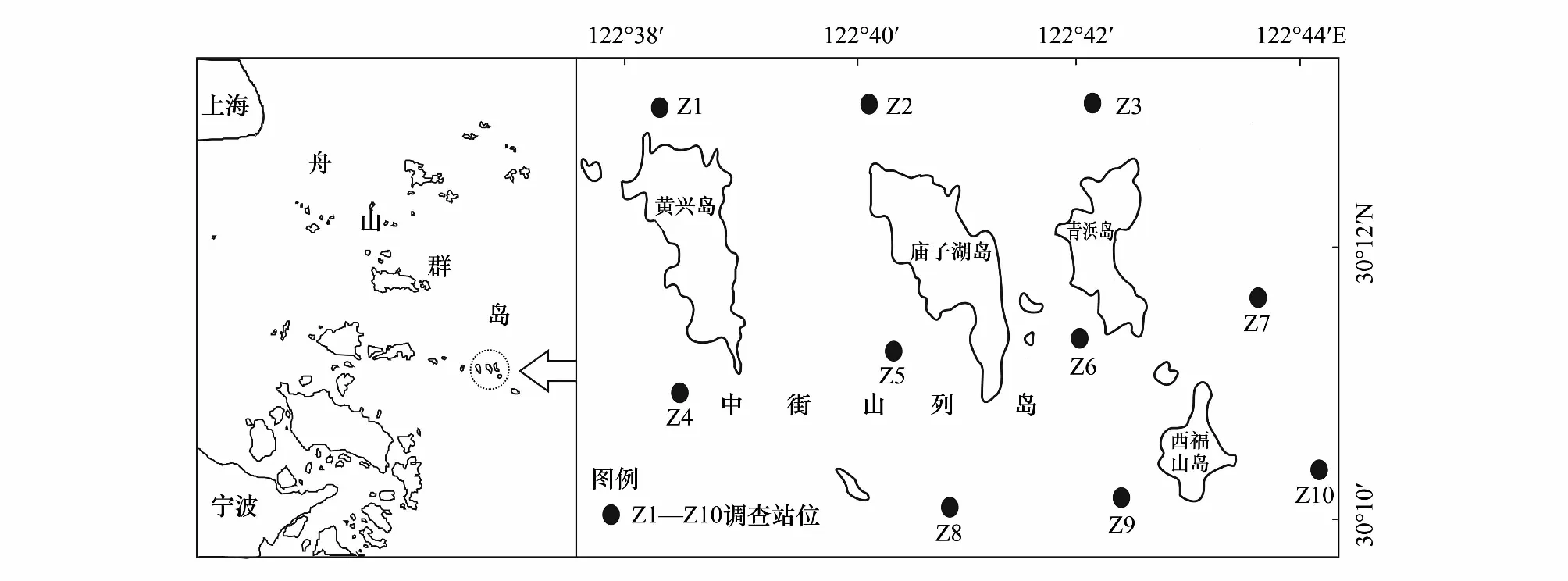
图1 中街山列岛海域鱼卵仔稚鱼调查站位Fig.1 Sampling station of ichthyoplankton in Zhongjieshan Islands
1.2 数据处理与分析方法
1.2.1生态优势度
鱼卵、仔稚鱼以每网实际数量(粒、尾) 来计算其密度,单位为粒/网或尾/网。
生态优势度利用Pianka[12]相对重要性指数(Index of Relative Importance,简称IRI) 计算,公式:
IRI=N%×F%×10000
式中,N%为某一种类鱼卵或仔稚鱼数量占采获鱼卵或仔稚鱼总量的百分比;F%为某一种类鱼卵或仔稚鱼出现的站位数占总调查站位数的百分比,IRI值大于1000为优势种,1000—100为重要种。
式中,IRIi为第i种鱼卵或仔稚鱼的相对重要性指数。
1.2.2物种多样性指数
香农-威纳指数(H′)计算公式
丰富度(D)计算公式:
D=(S-1)/log2N
均匀度(J′)计算公式:
1.2.3群落结构分析
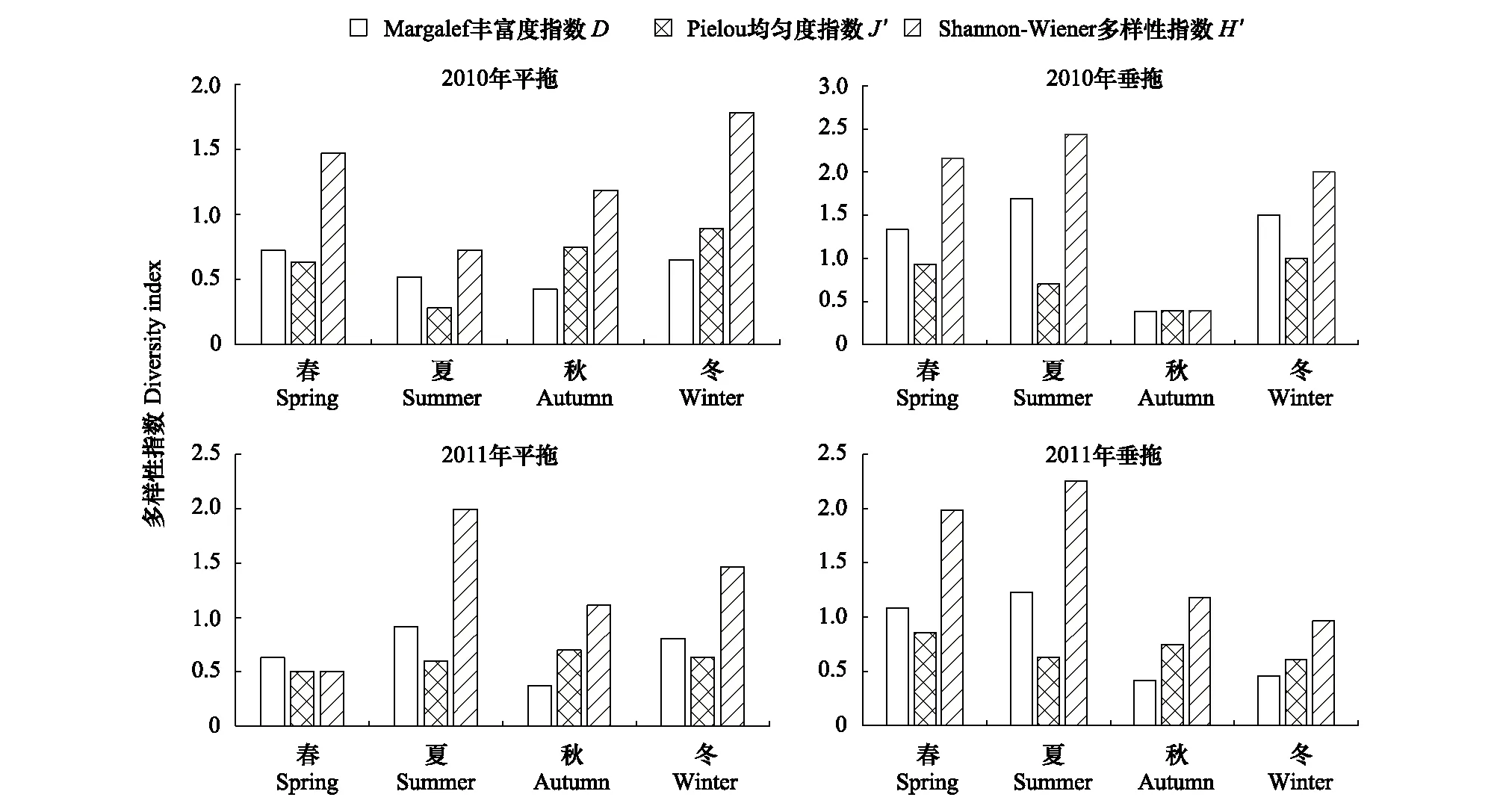
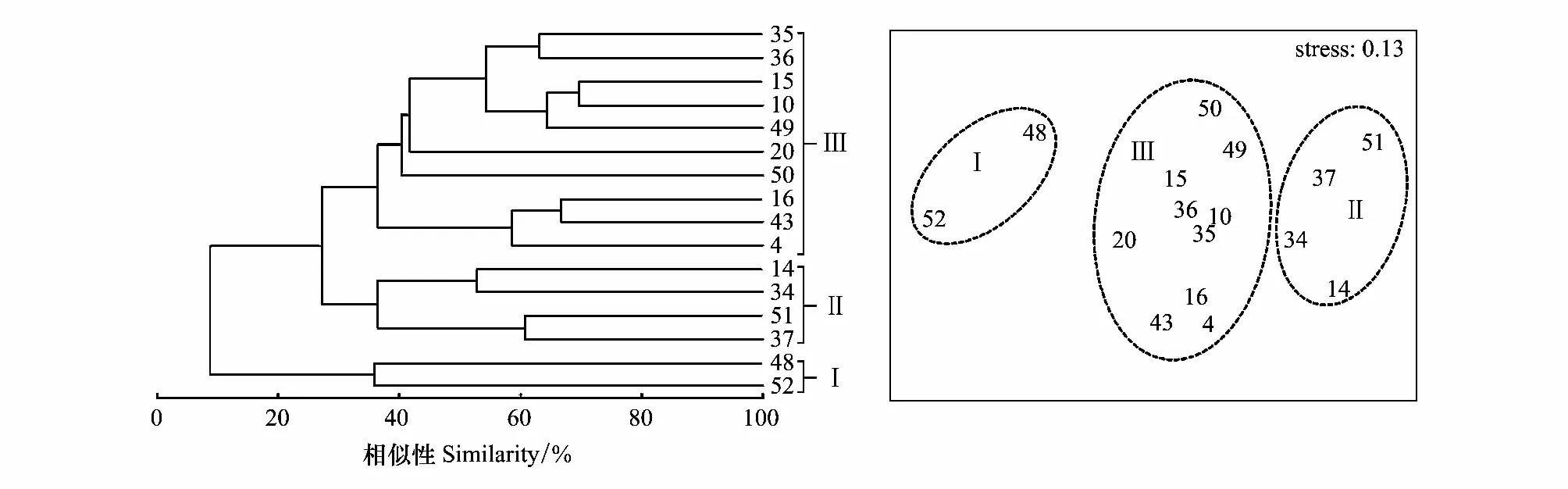
用Primer 5.0进行聚类分析(Cluster)和非度量多维标度排序(non-metric multi-dimensional scaling, NMDS)分析群落结构特征,对鱼卵、仔稚鱼数据采用对数转化[lg (x+1)]并进行相关分析。检验NMDS分析结果的优劣,用胁强系数(Stress)来衡量,当0.1 1.2.4环境因子分析 用Canoco 5.0对鱼类浮游生物与环境因子之间的关系进行分析及排序图绘制,以揭示不同环境因子对鱼卵、仔稚鱼的影响。对物种数据进行除趋势对应分析(DCA),根据每个轴的梯度长度(LGA) 选择适宜的排序方法。当 LGA<3 时,选择冗余分析(RDA);当 LGA>4 时,则可运用典范对应分析(CCA);当 3 2010年平拖和垂拖共采集鱼卵、仔稚鱼29种(不包括2个未定名物种),隶属于6目13科,有1种鉴定到目,7种鉴定到科,3种鉴定到属。其中,鲈形目种类最多,为5科10种,占占已鉴定出鱼卵、仔稚鱼种类数的34.48%;灯笼鱼目次之,为2科6种,占占已鉴定出鱼卵、仔稚鱼种类数的20.69%;鲉形目最少仅为1科1种,占已鉴定出鱼卵、仔稚鱼种类数的3.45%。平拖共采集鱼卵、仔稚鱼14种(不包括2个未定名物种),隶属于6目10科,4种鉴定到科,2种鉴定到属;垂拖共采集鱼卵、仔稚鱼21种,隶属于5目10科,有1种鉴定到目,4种鉴定到科,2种鉴定到属。 2011年平拖和垂拖共采集鱼卵、仔稚鱼25种(不包括6个未定名物种),隶属于6目14科,有1种鉴定到目,6种鉴定到科,1种鉴定到属。其中,鲈形目种类最多,为5科9种,占已鉴定出鱼卵、仔稚鱼种类数的36%;鲽形目次之,为2科5种,占已鉴定出鱼卵、仔稚鱼种类数的20%;月鱼目最少为1科1种,占已鉴定出鱼卵、仔稚鱼种类数的4%。平拖共采集鱼卵、仔稚鱼15种(不包括3个未定名物种),隶属于6目12科,有1种鉴定到目,4种鉴定到科,1种鉴定到属;垂拖共采集鱼卵、仔稚鱼17种(不包括3个未定名物种),隶属于5目11科,有3种鉴定到科(表1)。 表1 2010年和2011年中街山列岛海域鱼卵、仔稚鱼的种类组成 2010年平拖鱼卵仔稚鱼优势种为短吻红舌鳎(Cynoglossusjoyneri)(IRI%为81.54%),重要种为皮氏叫姑鱼(Johniusbelengeri)(IRI%为11.51%);垂拖鱼卵仔稚鱼重要种为鲉形目未定种1 (Scorpaeniformes sp.1)(IRI%为40%),扁舵鰹(Auxisthazard)(IRI%为38.10%)。 2011年平拖鱼卵仔稚鱼重要种为短吻红舌鳎(IRI%为46.12%)、鰏属未定种(Leiognathusssp.)(IRI%为32.99%)、龙头鱼(Harpodonnehereus)(IRI%为7.89%)和鳀(Engraulisjaponicus)(IRI%为6.59%);垂拖鱼卵仔稚鱼重要种为短吻红舌鳎(IRI%为52.12%)、龙头鱼(IRI%为23.26%)和中华小公鱼(Stolephoruschinensis)(IRI%为14.12%)。 表2 2010年和2011年中街山列岛海域鱼卵仔稚鱼优势种和重要种组成 从图2看出,2010年平拖丰富度指数(D)春季最高,而2011年夏季最高;2010年平拖均匀度指数(J′)冬季最高,2011年则是秋季最高;2010年平拖多样性指数(H′)冬季最高,2011年多样性指数(H′)夏季为最高值。 图2 2010年和2011年中街山列岛海域鱼卵、仔稚鱼的物种多样性指数Fig.2 Species diversity index of ichthyoplankton in Zhongjieshan Islands in 2010 and 2011 2010年和2011年垂拖丰富度指数(D)均是夏季最高;2010年垂拖均匀度指数(J′)冬季最高,2011年则是春季最高;2010年和2011年垂拖多样性指数(H′)均是夏季最高。 对2010—2011年中街山列岛海域平拖鱼卵仔稚鱼的16种优势种和重要种进行Bray-Curtis相似性指数分析,结果显示在35.81%的相似性指数水平上,可将16种鱼卵仔稚鱼优势种和重要种划分为3个类群,群落Ⅰ包含银鱼属未定种(Salanxsp.)和未定种4;群落Ⅱ包含鮻(Lizahaematocheilus)和褐菖鲉(Sebastiscusmarmoratus)等4种;群落Ⅲ包含斑头六线鱼(Agrammusagrammus)、龙头鱼和鮋科未定种(Scorpaenidae sp.)等10种(表1,图3)。ANOSIM分析结果GlobalR=0.782,P=0.001<0.01,表明3个类群间存在显著差异。 图3 2010—2011年中街山列岛海域平拖鱼卵仔稚鱼优势种和重要种聚类图和NMDS排序图(图中编号对应的种类名称见表1)Fig.3 Dominant and important species of ichthyoplankton in Zhongjieshan Islands demonstrated by clustering and NMDS ordination in 2010 and 2011 from horizontal tows (the species names corresponding to the numbers in the figure are shown in Tab. 1)NMDS: 非度量多维标度排序Non-metric multi-dimensional scaling 对2010—2011年中街山列岛海域垂拖鱼卵仔稚鱼的23种优势种和重要种进行Bray-Curtis相似性指数分析,结果显示在24.8%的相似性指数水平上,可将23种鱼卵仔稚鱼的优势种和重要种划分为4个类群,群落Ⅰ包含大泷六线鱼(Hexagrammosotakii)和白姑鱼(Argyrosomusargentatus)等6种,群落Ⅱ包含皮氏叫姑鱼(Johnius belengeri)和褐菖鲉等6种; 美肩鳃鳚(Omobranchuselegans)、拉氏狼牙虾虎鱼(Odontamblyopuslacepedii)、中华小公鱼和龙头鱼,群落Ⅲ包含虾虎鱼科未定种1 (Gobiidae sp.1),群落Ⅳ包含鯷和棘头梅童鱼(Collichthyslucidus)等10种(表1,图4)。ANOSIM分析结果Global R=0.789,P=0.001<0.01,表明4个类群间存在显著差异。 图4 2010—2011年中街山列岛海域垂拖鱼卵仔稚鱼优势种和重要种聚类图和NMDS排序图(图中编号对应的种类名称见表1)Fig.4 Dominant and important species of ichthyoplankton in Zhongjieshan Islands demonstrated by clustering and NMDS ordination in 2010 and 2011 from vertical tows (the species names corresponding to the numbers in the figure are shown in Tab. 1) 鱼卵群落相似性分析结果显示(表3),2010年和2011年的组内相似性分别为45.82%、50.32%,2010年主要特征种为短吻红舌鳎、皮氏叫姑鱼和鱚科未定种(Sillaginidaesp.)等,2011年主要特征种为短吻红舌鳎、中华小公鱼和龙头鱼等。 仔稚鱼群落相似性分析结果显示(表3),2010年和2011年的组内相似性分别为21.95%、36.72%,2010年主要特征种为鮻、褐菖鲉和大泷六线鱼,2011年的主要特征种为短吻红舌鳎、鯷和斑头六线鱼。 表3 2010—2011年中街山海域鱼卵仔稚鱼各群落内平均相似性贡献率之和占90%以上的特征种 群落相异性分析结果显示(表4),2010年和2011年鱼卵的相异性为74.53%,2010年和2011年仔稚鱼的相异性为94.54%。鱼卵的主要分歧种为鰏属未定种(Leiognathusssp.)、短吻红舌鳎和龙头鱼等;仔稚鱼的主要分歧种为斑头六线鱼、鮻和褐菖鲉等。 表4 2010—2011年中街山海域鱼卵仔稚鱼各群落间平均相异性贡献率之和占90%以上的分歧种 CCA分析结果显示,2010年对鱼卵、仔稚鱼群落结构影响较大的环境因子是温度和盐度(图2),第一轴CCA1,特征值=0.930,第二轴CCA2,特征值= 0.831物种—环境相关性(CCA1和CCA2 与环境因子相关性分别为0.996和0.965)。蒙特卡洛显著性检验结果显示,第一排序轴和所有排序轴均呈现为极显著差异(第一轴:F=0.2,P=0.034;所有轴:F=1.5,P=0.002)。两个排序轴共解释了主要鱼类组成21.33%的变异,其中,SSS及AS与第一排序轴呈较大正相关,而与SST呈较大负相关。舌鳎科、虾虎鱼科等鱼卵、仔稚鱼受底盐影响较大,矛尾虾虎鱼、皮氏叫姑鱼等鱼卵、仔稚鱼受悬浮物影响较大。 图5 2010年和2011年年中街山列岛海域鱼卵、仔稚鱼和环境因子之间的关系Fig.5 The relationship between environmental factors and ichthyoplankton in Zhongjieshan Islands in 2010 and 2011 CCA: 典范对应分析Canonical correspondence analysis;SST: 表温Surface temperature;SSS: 表盐Surface salinity;BST: 底温Bottom temperature;BST: 底盐Bottom salinity;SPM: 悬浮物Suspended matter;Am: 氨氮Ammonia nitrogen;Nr: 硝酸盐Nitrate;Nt: 亚硝酸盐Nitrite;Ni: 活性磷酸盐Active phosphate;AS: 活性硅酸盐Active silicate 2011年对鱼卵、仔稚鱼群落结构影响较大的环境因子是悬浮物、温度和盐度(图2),第一轴CCA1,特征值=0.976,第二轴CCA2,特征值= 0.506物种—环境相关性(CCA1和CCA2 与环境因子相关性分别为0.990和0.894)。蒙特卡洛显著性检验结果显示,第一排序轴和所有排序轴均呈现为极显著差异(第一轴:F=0.4,P=0.035;所有轴:F=2.0,P=0.001)。两个排序轴共解释了主要鱼类组成22.80%的变异,其中,SSS及SPM与第一排序轴呈较大正相关,而与BST呈较大负相关。大泷六线鱼、舒氏海龙受悬浮物影响较大,鲉形目未定种、龙头鱼和虾虎鱼科未定种受底盐影响较大。 表5 2010年和2011年中街山列岛海域基于CCA分析的鱼卵、仔稚鱼与环境因子的关系 鱼卵和仔稚鱼是鱼类生命周期中的初级阶段,其数量变化是衡量鱼类亲体资源量的重要指标[1]。中街山列岛海域2010年夏季和2011年夏季采获的鱼卵和仔稚鱼的数量占两年采获总量的89.58%,其出现频率和平均密度均高于其他季节,这表明夏季为该海域的产卵盛期。调查发现,中街山列岛海域2010年鱼卵、仔稚鱼的主要种类有短吻红舌鳎、皮氏叫姑鱼和鲉形目未定种等,更替为2011年短吻红舌鳎、鰏属未定种和龙头鱼等,两年主要鱼类均以底层小型鱼类为主。中街山列岛海域是小黄鱼、带鱼和银鲳等传统经济鱼类的产卵、育幼与索饵场所,且每年渔汛期都有大量的经济鱼类到此洄游,历史上也是曼氏无针乌贼重要的产卵场[15]。而本研究中仅皮氏叫姑鱼、龙头鱼和鳀等小型经济鱼类占有一定比重,随着人类的过度商业捕捞,沿岸工业的快速发展以及气候的变化,影响到渔业资源的分布和丰度[16],大黄鱼、小黄鱼等大个体经济鱼类遭受着过度破坏[17],该海域鱼类正在向小型和低营养层次鱼种转化。短吻红舌鳎是为舟山海域鲽形目鱼类中最为重要的种类之一[18],为暖温带中小型底层鱼类,栖息于泥沙质海底,产卵时间在3月下旬至8月中旬,为两年共同的优势种,其中,2010年夏季平拖密度527.89ind./100m3,2011年夏季平拖密度40.09ind./100m3,远高于其他物种的密度,这可能是由于夏季为短吻红舌鳎的产卵盛季。 中街山列岛海域岛礁星罗棋布,同时台湾暖流从浙江沿岸经过[19]和大陆径流带来了丰富的营养物质,为恋礁性鱼类提供了良好的栖息环境。聚类和排序分析结果显示,2010—2011年平拖鱼卵仔稚鱼的优势种和重要种3个类群间存在显著差异(图3)。群落Ⅱ包括褐菖鲉和大泷六线鱼等,褐菖鲉属于暖水性底层鱼类,是典型的岛礁定居性鱼类,以小鱼、甲壳类和端足类为食[20];大泷六线鱼是近海底层岩礁鱼类,以小型鱼类、甲壳类和多毛类为食[21],它们的生活习性相似;群落Ⅲ包含斑头六线鱼、皮氏叫姑鱼等恋礁鱼类,短吻红舌鳎和鰏属未定种等底层鱼类,它们多栖息于近海底层。研究发现,该海域与枸杞岛沙地岩礁生境的优势种相一致[22]。 2010—2011年垂拖鱼卵仔稚鱼的优势种和重要种4个类群间存在极显著差异(图4),群落Ⅱ包括皮氏叫姑鱼、鮻和七星底灯鱼等,皮氏叫姑鱼是近海洄游性底层鱼类, 喜栖息于泥沙底质和岩礁附近海域,摄食对虾、底栖桡足类和多毛类等[23]。鮻为暖温性底层鱼类,以食浮游硅藻和桡足类为主;七星底灯鱼为中、底层小型鱼类,以桡足类或其他甲壳动物的幼体为食;它们有着相似的摄食习性和栖息环境。群落Ⅳ包含斑头六线鱼、短吻红舌鳎、棘头梅童鱼和龙头鱼等,大多为短距离洄游和定居种。通过对两年间平拖和垂拖鱼卵仔稚鱼的分析发现,该海域以恋礁的小型底层鱼类为主,这与汪洋等[24]对中街山列岛岩礁海域成鱼群落多样性的研究结果相似。 鱼卵群落相似性分析结果显示(表3),2010年主要特征种为短吻红舌鳎、皮氏叫姑鱼和鱚科未定种1等,2011年主要特征种为短吻红舌鳎、中华小公鱼和龙头鱼;仔稚鱼群落相似性分析结果显示(表3),2010年主要特征种为鮻、褐菖鲉和大泷六线鱼,2011年的主要特征种为短吻红舌鳎、鯷和斑头六线鱼。在鱼卵群落结构中,短吻红舌鳎、龙头鱼为两年共同种,且两种都为定居性鱼类;在仔稚鱼群落结构中,褐菖鲉、斑头六线鱼和鮻等在当年很多季节都有出现,其种类大多为定居性鱼类和恋礁性鱼类。分析发现,定居性鱼类和恋礁性鱼类对该海域岩礁生境周年依赖,因此年际间鱼卵、仔稚鱼主要特征种变化不大。 鱼类多样性和海洋环境密切相关,环境因子影响鱼类栖息地的偏好,塑造了不同的空间分布格局[25- 27],同时环境因子也影响着鱼类生长、存活及代谢等机能活动,鱼卵仔稚鱼和繁殖期的亲鱼对水温和盐度的变化特别敏感[28- 30]。中街山列岛海域2010年和2011年夏季的垂直拖网、2011年夏季水平拖网的鱼卵仔稚鱼群落的生物多样性指数最高,两年的鱼卵数量均为夏季最高,夏季海水中营养物质丰富,鱼类产卵索饵与水温、盐度和溶解氧等环境因素密不可分,适宜的温度促进鱼卵、仔稚鱼的代谢和运动,进而影响其生长发育[31]。 CCA排序图表明,温度和盐度对鱼卵、仔稚鱼群落结构影响较大,水温是影响鱼类生理、生化和生活史过程的重要环境因子,它在促使鱼类性腺发育和诱使鱼类产卵繁殖方面起着重要作用[32]。盐度影响鱼体渗透压调节的改变,并且还能引起鱼体与环境间物质交换与能量流动的改变,对鱼类的生长和摄食产生一定的影响[33]。鱼卵、仔稚鱼群落结构、空间等能在一定程度上反映水体的水质状况[34]。研究发现中街山列岛海域在2010、2011年海水水质富营养化相对严重[35],海域富营养化程度与活性磷酸盐含量密切相关,因此大部分鱼卵、仔稚鱼的分布离活性磷酸盐较远。邱东茹等[36]研究认为悬浮物多不利于靠视觉定位的凶猛性鱼类捕食,从而减轻了对摄食浮游生物和底栖生物的鱼类的捕食压力。大泷六线鱼、矛尾虾虎鱼和皮氏叫姑鱼均摄食底栖生物类、多毛类等,它们分布于悬浮物较高的区域,易于躲避凶猛性鱼类捕食。2 结果
2.1 种类组成

2.2 优势种及其季节变化

2.3 物种多样性指数
2.4 群落结构分析
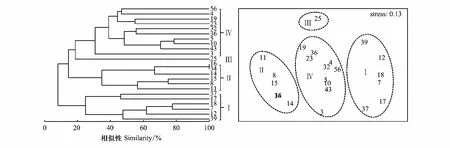
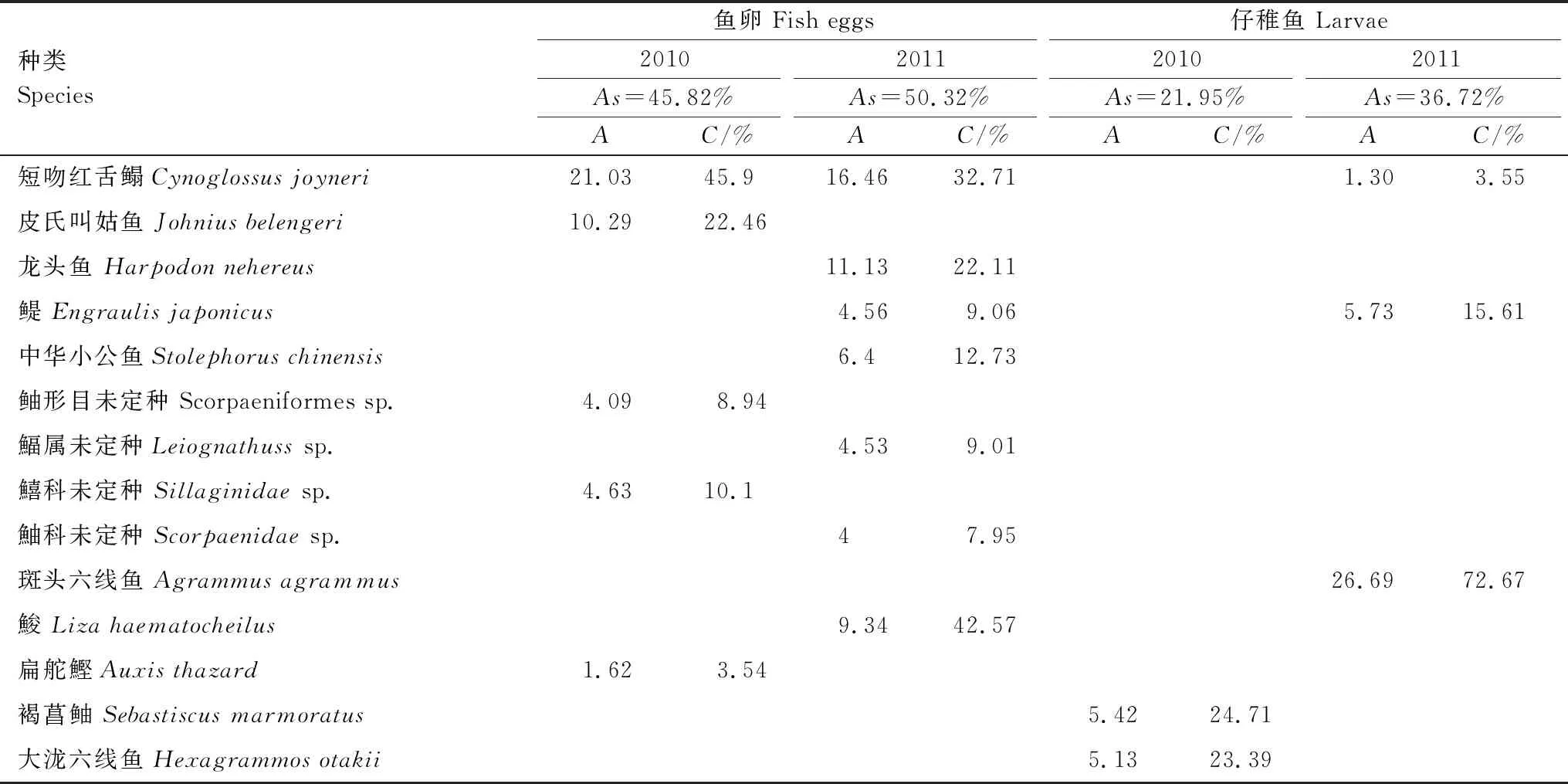
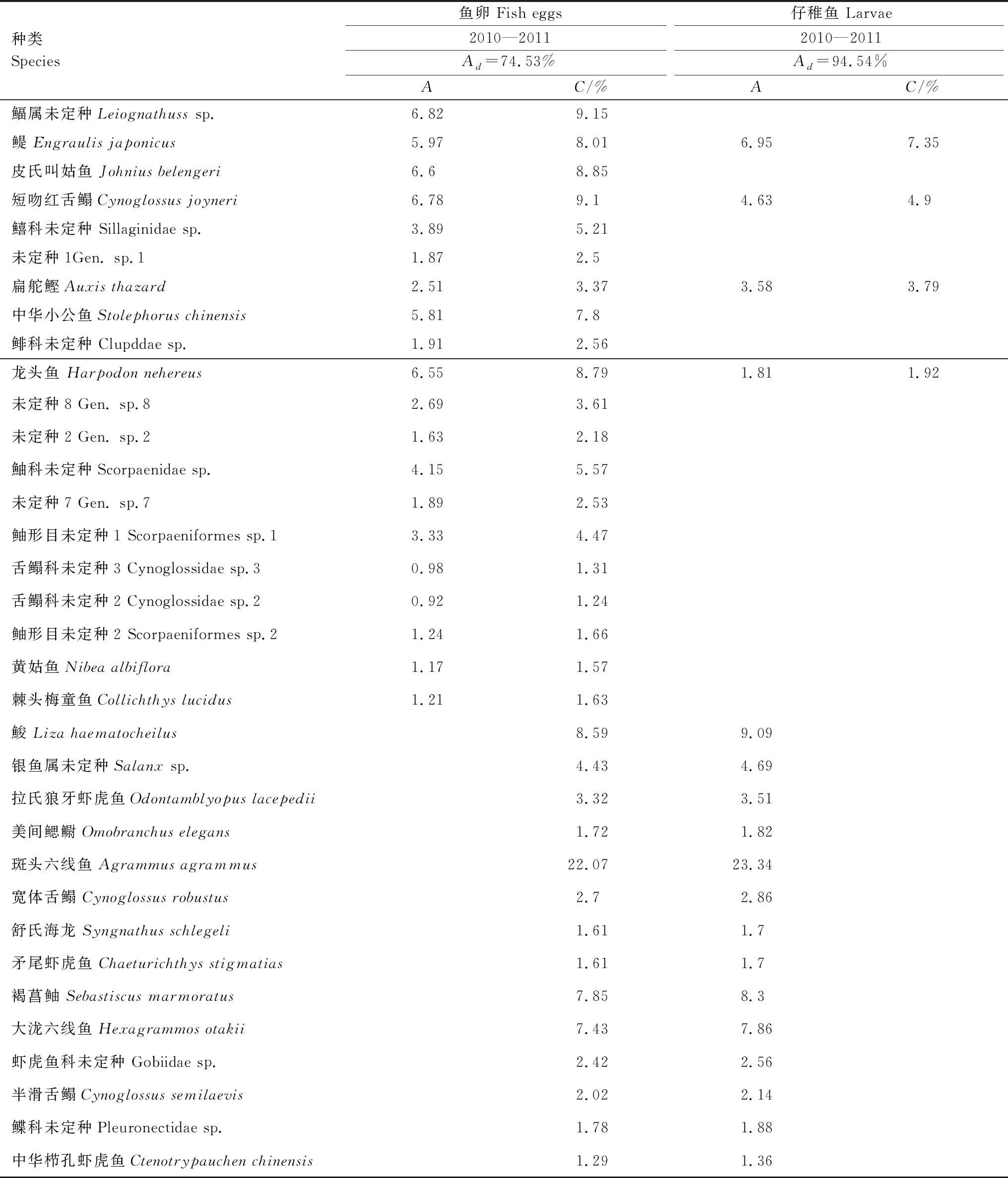
2.5 群落相似性分析
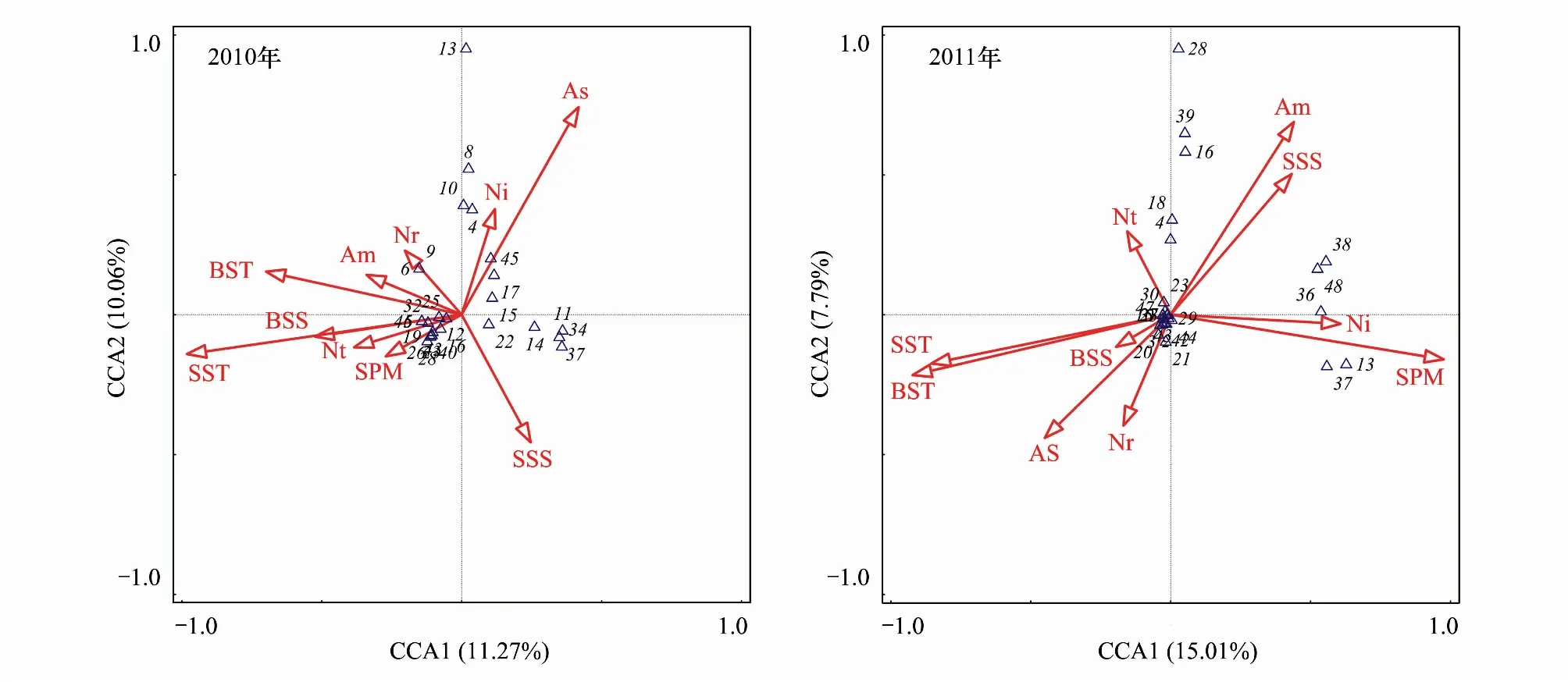
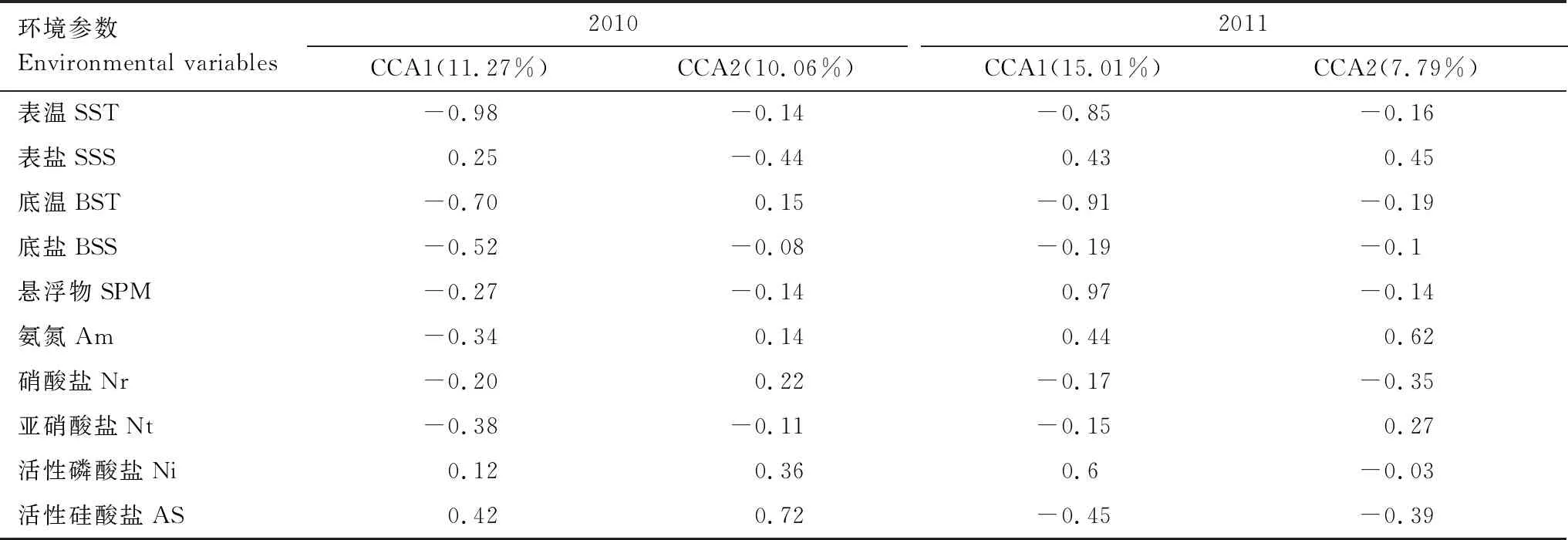
2.6 鱼卵仔稚鱼群落结构与环境因子的关系
3 讨论
3.1 鱼卵、仔稚鱼群落组成
3.2 鱼卵、仔稚鱼与环境因子的相关性
