HGGT基因与小麦赤霉病籽粒毒素积累的关系解析
2020-08-04吴迪朱素芹张语卉
吴迪 朱素芹 张语卉


摘要:赤霉病是我国小麦重要病害之一,禾谷镰刀菌是其主要致病因子。赤霉病菌在侵染小麦过程中会在籽粒中产生脱氧雪腐镰刀菌烯醇毒素(DON)。维生素E的抗氧化作用在一定程度上能降低DON的毒性,HGGT(尿黑酸牻牛儿基牻牛儿基转移酶)是维生素E生物合成的关键限速酶。为了揭示HGGT基因与小麦赤霉病及DON毒素积累的关系,本研究克隆HGGT基因的全长,并开发了基因特异标记,在31份小麦品种中进行标记分析,明确了该基因的分布。同时采用双花滴注法在扬花期对31份小麦品种进行赤霉病接种和抗性鉴定,成熟期利用LC-MS法测定籽粒中DON毒素含量。结果表明,HGGT基因与赤霉病抗性相关,携带HGGT基因的小麦品种病小穗率及籽粒毒素含量显著(P<0.05)低于不携带该基因的品种,说明HGGT基因可以用于改良小麦赤霉病抗性,并降低籽粒毒素含量。
关键词:小麦;赤霉病;DON毒素;HGGT基因;维生素E
中图分类号: S435.121.4+5 文献标志码: A 文章编号:1002-1302(2020)11-0096-04
收稿日期:2020-04-20
基金项目:国家自然科学基金(编号:31771772)。
作者简介:吴 迪(1992—),男,江苏扬州人,硕士研究生,主要从事小麦遗传育种研究。E-mail:956671930@qq.com。
通信作者:李 韬,博士,教授,主要从事小麦赤霉病遗传和种质创新研究。E-mail:taoli@yzu.edu.cn。 小麦赤霉病是由禾谷镰刀菌引起的一种麦类真菌病害。赤霉病抗性类型主要分为抗侵染、抗扩展、籽粒抗性和抗毒素积累。小麦赤霉病多发生在温暖多雨和气候湿润地区[1],近年来,我国小麦赤霉病发生区域由长江流域向黄淮流域不断北移[2],主要发生在长江中下游麦区和东北春麦区[3]。禾谷镰刀菌侵染小麦后造成穗部组织褐化坏死,籽粒干瘪,导致小麦产量和品质下降[4-5],造成严重的经济损失。小麦赤霉病的另一大危害是感病籽粒中积累的毒性次级代谢产物脱氧雪腐镰刀菌烯醇(DON)[6-7],人畜中毒后会出现胃部不适,恶心眩晕、头痛[8],故又称呕吐毒素,还会造成神经危害,抑制机体免疫反应[9]。小麦赤霉病不仅造成粮食减产,还在侵染过程中积累毒素,造成严重的食品安全隐患[10]。因此赤霉病引起的食品和饲料污染问题引起了人们的极大关注。我国卫生和标准管理部门规定,小麦和玉米中DON含量超过1 mg/kg、小麦赤霉病病粒含量超过4%时则禁止食用[11]。
维生素E是一种重要的抗氧化剂,可分为生育酚和生育三烯酚两大类[12]。小麦中的生育酚主要存在于胚中,生育三烯酚存在于果皮和胚乳中[13]。维生素E的生物合成途径包括:(1)亲水性头部的合成。(2)HGA的植基化。HGA分别在尿黑酸植基转移酶(HPT)和尿黑酸牻牛儿基牻牛儿基转移酶(HGGT)的催化下生成生育酚和生育三烯酚的前体物[14]。(3)MPBQ(2-甲基-6-植基-苯醌)和MGGBQ(2-甲基-6-牻牛儿牻牛儿基-苯醌)的甲基化。(4)环化反应[15]。HGGT基因定位于质体中,主要在小麦胚乳中表达。HGGT是维生素E生物合成的关键酶基因[16],Cahoon等于2003年第一次从大麦和水稻种子中克隆得到HGGT序列[17]。小麦籽粒积累DON毒素后,活性氧(ROS)含量提高,抗氧化能力下降,诱导机体发生氧化应激反应,最终导致机体和细胞的脂质过氧化和DNA氧化损伤[18]。维生素E是一种强自由基清除剂,能及时清除活性氧,防止脂質过氧化的产生[19]。在饲料中添加维生素E能降低DON的毒性作用,在膳食中使用维生素E能缓解DON引起的急性中毒[20],但维生素E对DON的降毒机制尚不清楚。HGGT是生育三烯酚合成的第一个限速关键酶,也是维生素E合成的第一个分支点[21]。本试验的主要目的是在31份小麦品种中扩增HGGT基因,接种赤霉菌鉴定病小穗率,并测定籽粒中的毒素含量,解析HGGT基因与小麦赤霉病抗性、病小穗率及毒素含量的关系。
1 材料与方法
1.1 试验材料
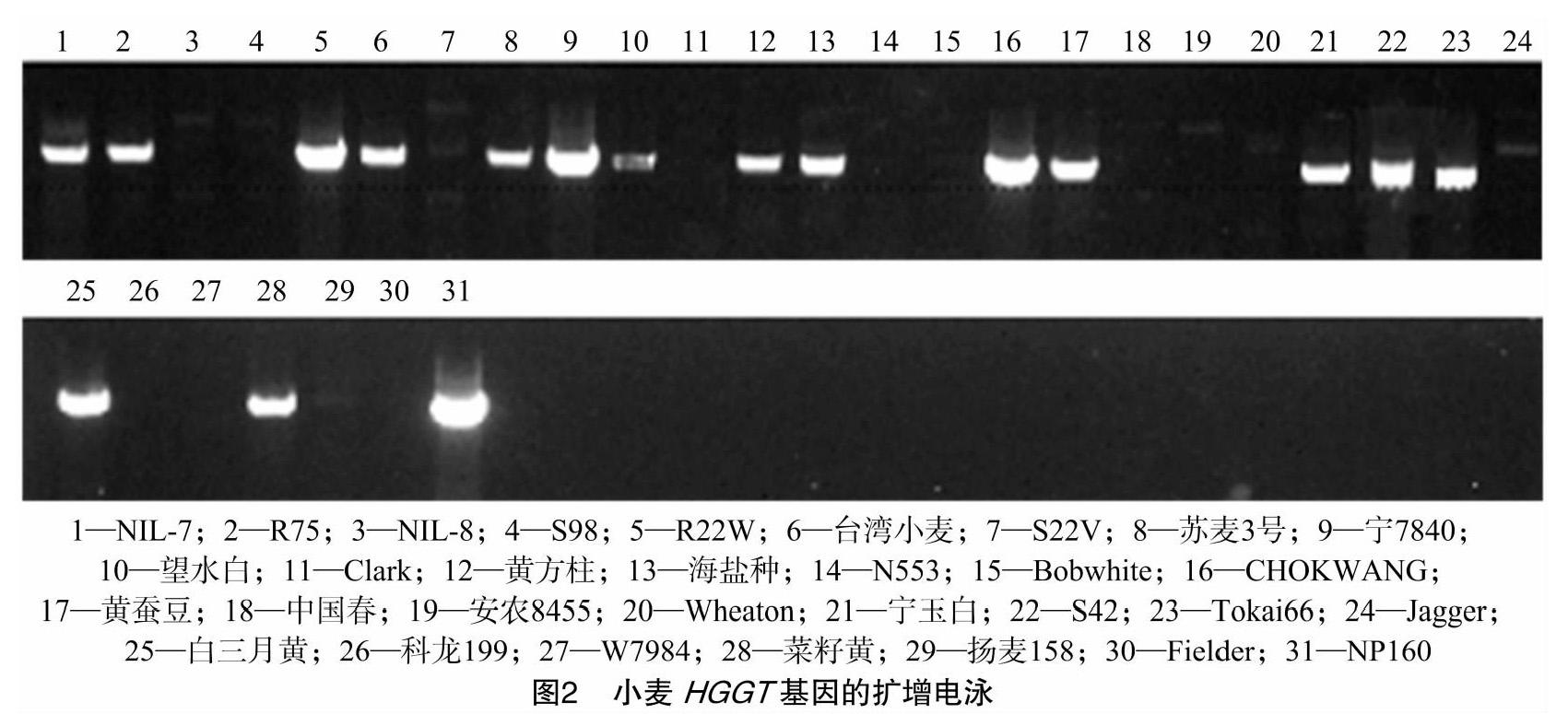
31份小麦材料为:NIL-7、R75、NIL-8、S98、R22W、台湾小麦、S22V、苏麦3号、宁7840、望水白、Clark、黄方柱、海盐种、N553、S42、CHOKWANG、黄蚕豆、中国春、安农8455、Wheaton、宁玉白、Bobwhite、Tokai66、Jagger、白三月黄、科龙199、W7984、菜籽黄、扬麦158、Fielder、NP160。所有材料均于2018年10月种植于扬州大学试验田,每个品种播种6行,每行15株。
1.2 小麦基因组DNA提取
苗期取31份材料的嫩叶,采用CTAB法[22]提取各材料的基因组DNA。用超微量分光光度计检测出的DNA浓度均在600 ng/μL左右,D260 nm/D280 nm处于1.7~2.0之间。
1.3 HGGT基因引物设计及扩增
通过GenBank网站找到HGGT基因的序列信息,利用Primer Premier 5.0软件设计HGGT基因的特异扩增引物(由华大基因公司合成)。
以上述提取获得的小麦基因组DNA为模板,Seq400.6-2F/Seq400.6-R为引物进行PCR扩增,以鉴定HGGT基因在小麦赤霉病不同抗性品种中的分布。PCR反应采用50 μL体系:上下游引物各1 μL,DNA模板10 μL,2×Gflex Buffer 25 μL,ddH2O 12 μL,TKs Gflex酶1 μL。PCR反应在Thermal Cycler C1000 Touch PCR仪上进行,程序如下:95 ℃预变性2 min;95 ℃变性30 s,55 ℃退火 30 s,72 ℃延伸210 s,37个循环;72 ℃延伸5 min,4 ℃ 保存。取5 μL PCR产物进行1%的琼脂糖凝胶电泳检测带型,以确定其基因型。
1.4 PDA培养基的配制
称取40 g马铃薯琼脂糖(PDA)培养基于1 L烧杯中,加入1 L的ddH2O,用玻璃棒搅拌均匀,置于微波炉中加热15 min完全溶解至透明状,双层纱布过滤至锥形瓶中,高温高压灭菌锅121 ℃灭菌 20 min,待温度降至55 ℃时,于无菌操作台下倾注平板,配制25 mL的等量PDA培养基于9 cm直径的无菌培养皿中,待PDA培养基完全凝固后置于 25 ℃ 培养箱贮藏备用。
1.5 试验用菌株及其菌液制备
本试验所用禾谷镰刀菌菌株为F1312(江苏省农业科学院陈怀谷研究员提供)。利用灭菌后的接种环挑取纯菌株斜面上的禾谷镰刀菌边缘菌体,接种于PDA培养基上,置于25 ℃恒温培养5~7 d,用灭菌后直径7 mm的圆形打孔器从经过活化的禾谷镰刀菌菌株PDA培养基上,取8块菌块放入80 mL经过灭菌的绿豆汤培养基,然后置于摇床上,在 180 r/min、26 ℃下振摇72~120 h,吸取1 μL孢子液,滴到血球计数板上,置于显微镜下测定孢子浓度,然后稀释或浓缩到接种需要的浓度(1×105个/mL)。
1.6 赤霉病接种及鉴定
采用双花滴注法(bilateral florets inoculation method,BFI),在小麦扬花期将制备好的菌液注射到倒5小穗双侧小花的内外颖之间,每行接种10个穗子,记载接种日期,接种后21 d鉴定感病小穗数,计算病小穗率(proportion of symptomatic spikelets,PSS):PSS=感病小穗数/总小穗数。
1.7 籽粒中DON含量的测定
1.7.1 实验仪器 液质联用仪(TSQ-Vantage,Thermo Fisher SCIENTIFIC,美国)。
1.7.2 标准液 制备毒素原液DON(FERMENTEK,以色列)用10%乙腈水配制成 100 μg/L 混标。
1.7.3 样品前处理 前处理方法参照靳梦瞳等的方法[23]。利用磨样机将麦粒磨碎,称取2 g样品于50 mL离心管中,加入10 mL乙腈/水溶液,振荡后超声30 min,离心5 min后吸2 mL上清于新的 10 mL 离心管,加入150 mg无水硫酸镁,振荡吸上清至新的10 mL离心管。加入1 mL正己烷脱脂,离心去除正己烷层,氮气吹干剩余上清液,最后乙腈/水溶液定容至1 mL。涡旋过0.22 μm尼龙滤膜后,进样LC-MS分析。
1.7.4 色谱及质谱条件 色谱及质谱条件参照靳梦瞳等的方法[23],略有改进。主要参数如下:
色谱条件:色谱柱选用ACQUITY UPLC HSS T3(2.1×100 mm,1.8 μm),柱温40 ℃,样品温度 5 ℃。流速0.3 mL/min,进样体积为5 μL。采用流动相A(甲醇)和流动相B(乙酸铵)组成的流动相进行梯度洗脱。洗脱程序:0 min 20% A-80% B,1 min 20% A-80% B、5.5 min 90% A-10% B,65 min 90% A-10% B、7 min 20% A-80% B。
质谱条件:选择反应监测模式(MRM)检测。曲线脱溶剂管(CDL)温度250 ℃,雾化气体和干燥气体均为氮气,流速分别为3.0、15 L/min。碰撞气为高纯氩气,碰撞诱导解离压力为230 kPa。
2 结果与分析
2.1 HGGT基因的扩增
本研究通过BAC克隆(登录号:KU641029)获得HGGT基因,HGGT基因序列包含2個外显子和1个内含子。其编码区包含273个碱基,编码80个氨基酸。转录起始位点位于402 532 nt,第1外显子分布在401 715~401 828 nt,共114个碱基,第2外显子分布在400 820~400 975 nt,共156个碱基,PolyA信号位点位于400 620 nt(图1)。
为了鉴定HGGT基因在小麦赤霉病不同抗性品种中的分布,根据HGGT基因的序列设计特异性引物,进行了PCR扩增。通过1%的琼脂糖凝胶电泳检测PCR产物的带型,结果如图2所示:17份携带HGGT基因的品种都能扩增2 000 bp目的条带,其余14份品种中没有扩增条带。
2.2 不同小麦材料赤霉病病小穗率及籽粒DON含量
测定不同小麦材料的病小穗率及籽粒中的DON含量,结果(表1)表明,携带HGGT基因的17份材料高抗赤霉病,不携带HGGT基因的14份材料高感赤霉病。
2.3 2种基因型下的病小穗率及籽粒DON含量的差异
方差分析结果表明,携带HGGT基因与不携带HGGT基因的品种之间病小穗率存在差异,P=000<0.05;携带HGGT基因与不携带HGGT基因的品种之间的籽粒DON存在差异,P=0.00<0.05(表2)。说明携带HGGT基因与不携带HGGT基因的品种之间的病小穗率和籽粒毒素含量均存在差异。
3 讨论与结论
本试验根据HGGT基因序列,设计特异性标记,在31份不同赤霉病抗性小麦品种中利用基因扩增技术扩增目的基因,确定HGGT在不同赤霉病抗性品种中的分布。在扬花期进行赤霉病接种抗性鉴定,后续测定籽粒中积累的DON毒素含量,结果表明,HGGT基因只在高抗赤霉病品种中扩增出,在感病品种中未扩增出目的条带,且携带HGGT基因的小麦品种病小穗率及籽粒毒素含量较低,与不携带HGGT基因的小麦品种病小穗率及籽粒毒素含量存在显著差异(P<0.05),这也从侧面说明了籽粒中DON毒素积累的前提是赤霉菌在穗部侵染。说明HGGT基因与赤霉病扩展抗性有关,在一定程度上阻止病原菌在穗部的扩展并降低籽粒中积累的DON毒素含量,可以用于改良小麦赤霉病抗性。HGGT基因是维生素E生物合成途径的一个关键酶基因,我们推测维生素E对赤霉病的降毒效应,很有可能是受HGGT基因调控的作用,后续将进一步研究HGGT基因相关的功能以及HGGT基因对赤霉
病的抗性机制。赤霉菌在扬花期和抽穗期都会侵染小麦,会在籽粒中积累DON毒素。本研究结果证明了携带HGGT基因的小麦品种病小穗率及籽粒毒素积累量较低,说明HGGT基因具有赤霉病抗性作用,丰富了HGGT基因功能的认识,为小麦赤霉病病小穗率及DON毒素防控提供新的方向,对解析扩展抗性的机制和DON毒素方面的研究具有重要的参考价值。
参考文献:
[1]马跃亭. 小麦籽粒DON含量影响因素相关性分析及氨基酸对赤霉菌产毒的影响[D]. 镇江:江苏科技大学,2019:1-57.
[2]范三红,胡小平. 小麦赤霉菌毒素合成机制及检测技术研究进展[J]. 麦类作物学报,2018,38(3):348-357.
[3]程顺和,郭文善,王龙俊,等. 中国南方小麦[M]. 南京:江苏科学技术出版社,2012:281-282.
[4]Dexter J E,Clear R M,Preston K R. Fusarium head blight:effect on the milling and baking of some Canadian wheats[J]. Cereal Chemistry,1996,73(6):695-701.
[5]Dexter J E,Marchylo B A,Clear R M,et al. Effect of Fusarium head blight on semolina milling and pasta-making quality of durum wheat[J]. Cereal Chemistry,1997,74(5):519-525.
[6]Jiang C,Zhang C K,Wu C L,et al. Tri6 and Tri10 play different roles in the regulation of deoxynivalenol(DON) production by cAMPsignalling in Fusarium graminearum[J]. Environmental Microbiology,2016,18(11):3689-3701.
[7]Li T,Bai G,Wu S,et al. Quantitative trait loci for resistance to Fusarium head blight in a Chinese wheat landrace Haiyanzhong[J]. Theoretical and Applied Genetics,2011,122(8):1497-1502.
[8]徐存宽,黄 伟,孙宝胜. 呕吐毒素在小麦正常粒和赤霉病粒中的分布情况研究[J]. 粮食与油脂,2019,32(1):84-86.
[9]Woloshuk C P,Won-Bo S. Aflatoxins,fumonisins,and trichothecenes:a convergence of knowledge[J]. Fems Microbiology Reviews,2013,37(1):94-109.
[10]刘易科,佟汉文,朱展望,等. 小麦赤霉病抗性改良研究进展[J]. 麦类作物学报,2016,36(1):51-57.
[11]张爱民,阳文龙,李 欣,等. 小麦抗赤霉病研究现状与展望[J]. 遗传,2018,40(10):858-873.
[12]柴玉琼. 利用维生素E生物合成相关基因调控大麦生育酚含量及其组成[D]. 杭州:浙江大学,2015:1-80.
[13]张玉红,陈建澍,曾章慧,等. 大麦籽粒生育酚和生育三烯酚的合成与分布特性[J]. 植物生理学报,2015(4):181-185.
[14]Rohmer M. Mevalonate-independent methylerythritol phosphate pathway for isoprenoid biosynthesis.Elucidation and distribution[J]. Pure and Applied Chemistry,2003,75(2/3):375-387.
[15]Babura S R,Abdullah S N A,Khaza A H. Advances in genetic improvement for tocotrienol production:a review[J]. Journal of Nutritional Science &Vitaminology,2017,63(4):215-221.
[16]Yang W,Cahoon R E,Hunter S C,et al. Vitamin E biosynthesis:functional characterization of the monocot homogentisate geranylgeranyl transferase[J]. The Plant Journal,2011,65(2):206-217.
[17]Cahoon E B,Hall S E,RippK G,et al. Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content[J]. Nature Biotechnology,2003,21(9):1082-1087.
[18]王 旭,黄德玉,吴庆华,等. 真菌毒素引起的氧化应激及其毒理学意义[J]. 生态毒理学报,2015,10(6):62-70.
[19]杨 巍. 脱氧雪腐镰刀菌烯醇对人外周血淋巴细胞的遗传毒性研究[D]. 武汉:华中科技大学,2012:1-144.
[20]Rizzo A F,Atroshi F,Ahotupa M,et al. Protective effect of antioxidants against free radical-mediated lipid peroxidation induced by DON or T-2 toxin[J]. ZentralblVeterinarmed A,1994,41(1-10):81-90.
[21]FarreG,Sudhakar D,Naqvi S,et al. Transgenic rice grains expressing a heterologous β-hydroxyphenylpyruvate dioxygenase shift tocopherol synthesis from the γ to the α-isoform without increasing absolute tocopherol levels[J]. Transgenic Research,2012,21(5):1093-1097.
[22]施 璇. 小麦类过敏反应突变体抗病性研究及氮依赖性类过敏反应基因的定位[D]. 扬州:扬州大学,2018:1-72.
[23]靳梦曈,王建华,林善海,等. 室温下麦粒中脱氧雪腐镰刀菌烯醇(DON)及其隐蔽型(D3G)毒素累積分析[J]. 食品工业科技,2015,36(17):132-136.雷娇娇,田 力,袁 伟,等. 贵阳花溪久安茶树炭疽病病原菌(Colletotrichun gloeosporioides)的分离鉴定及生物学特性[J]. 江苏农业科学,2020,48(11):100-105.
