青藏高原高寒草甸退化对土壤氮素转化微生物基因的影响
2020-08-04王向涛廖李荣3喻岚辉3张宪洲
王向涛, 张 超, 廖李荣3,, 王 杰, 喻岚辉3,, 张宪洲
(1.西藏农牧学院 动物科学学院, 西藏 林芝 860000; 2.中国科学院 地理科学与资源研究所,生态系统网络观测与模拟重点实验室, 拉萨高原生态试验站, 北京 100101; 3.中国科学院大学, 北京 100049;4.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 5.中国科学院大学 资源与环境学院, 北京 100190)
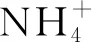
目前,对于青藏高原高寒草甸生态系统已开展了大量研究,如植物群落的演替[7]、土壤养分[8]以及微生物量[9]、酶活性[10]的变化,而对于土壤参与氮素转化的微生物研究相对较少,而这是深刻认识草地退化的关键科学问题。鉴于此,本研究以青藏高原不同退化程度的高寒草甸为对象,分析退化过程中土壤氮素转化微生物基因丰度的变化,明确其影响因素,为退化高寒草甸的科学治理和人工恢复提供参考依据。
1 材料与方法
1.1 研究区概况
研究区位于西藏自治区拉萨市当雄县草原站(91°05′E,30°51′N),海拔高度4 330 m,年均温为1.3 ℃,最高月(7月)平均温度为10.7 ℃,最低月(1月)平均温度为10.4 ℃。年平均降水量为476 mm,集中在6—8月之间。草地类型为高寒草甸,优势种为高山嵩草(Kobresiapygmaea),常见伴生种有窄叶苔草(Carexmontis-everestii)、丝颖针茅(Stipacapillacea)、二裂叶委陵菜(Potentillabifurca)和钉柱委陵菜(Potentillasaundersiana)等。土壤类型为典型的高山草甸土。
1.2 样品采集
根据草地植物种类及盖度的差异,参照Xue等[11]对天然草地退化等级划分标准,设定为未退化(non-degraded, ND),轻度退化(light degraded, LD),中度退化(moderate degraded, MD)以及重度退化(severe degraded, SD)4种退化草地处理。每个小区面积为40 m×60 m,每个处理5个重复。于2018年8月采集样品,采用蛇形取样法,每个小区沿150 m的样条建立了5个等间距的采样点(0.5 m×0.5 m),以内径为3.5 cm 的土钻,采集0—20 cm 土样,每样地取样5~8个,混为1个土样。土壤样本去除枯落物及植物根系后,分为两个部分,一部分过筛风干进行理化性质的测定分析,另一部分保存在-80 ℃用于进行DNA的提取。
1.3 土壤理化性质测定
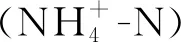
1.4 土壤微生物功能基因丰度测定
微生物功能基因丰度采用定量PCR测定。本研究中主要测定了参与土壤氮素转化的7个基因,包括nifH,amoA-AOA,amoA-AOB,narG,nirK,nirS以及nosZ,其功能见表1,测定方法参考廖李容等[13],即取0.25 g冻干样品,按照UltraClean soil DNA Extraction Kit试剂盒的说明书,提取土壤总DNA,目的基因引物序列见表1,扩增程序为98 ℃,持续60 s;30个循环,98 ℃,10 s;50 ℃,30 s;72 ℃,30 s;在72 ℃下保持5 min,每个样品重复3次,使用ABI 7500实时PCR系统测定浓度。

表1 氮转化微生物功能基因PCR扩增引物及对应功能[13]
1.5 数据处理
采用单因素方差分析对不同退化程度草地的土壤理化性质及功能基因进行差异分析,并使用最小显著性差异法对不同退化草地之间的差异进行多重比较,显著水平为0.05。采用Pearson法和冗余分析 (RDA),分析氮转化基因与土壤性质之间的关系。所有统计分析采用R的vegan包,利用SigmaPlot 12.0作图。图中数据为平均值±标准差。
2 结果与分析
2.1 土壤理化性质
高寒草甸退化显著影响土壤理化性质(见表2)。与未退化草甸相比,退化草甸的土壤有机碳、全氮、硝态氮和铵态氮含量均显著降低(p<0.05),降幅分别达41.9%~67.5%,26.8%~56.2%,44.7%~76.6%和36.0%~58.9%。轻度退化草甸的速效磷及速效钾含量与未退化草甸相比无显著差异(p>0.05),显著高于中度和重度退化草甸(p<0.05)。土壤pH值变化为6.43~6.90,且随退化程度的增高而逐渐增加。退化降低了土壤水分,其含量随退化程度的增高而逐渐下降,与未退化草甸相比,重度退化草甸的水分含量降低了33.6%。
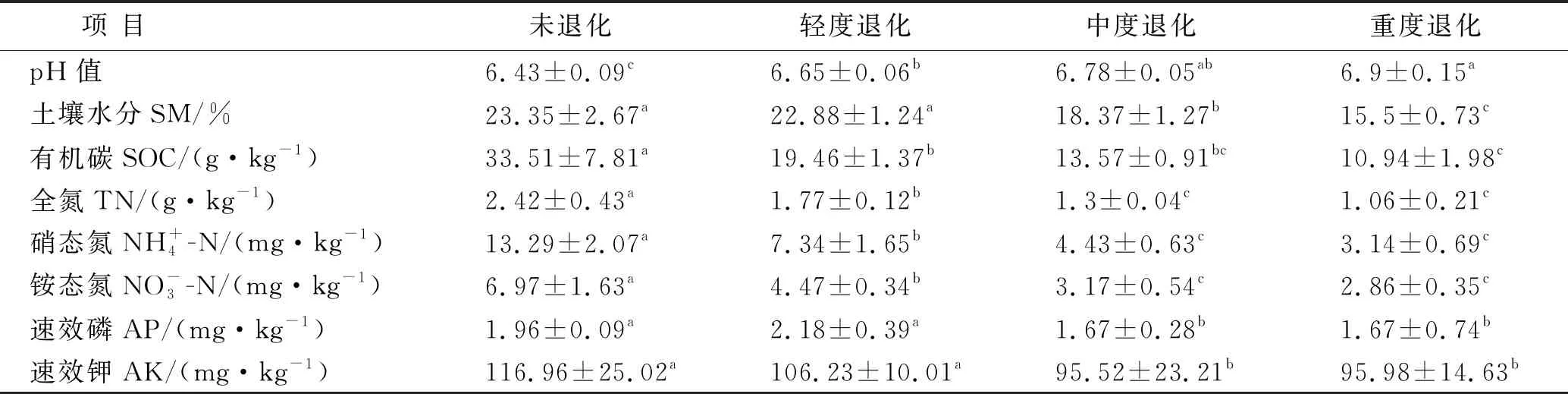
表2 不同退化草地土壤理化性质
2.2 土壤氮素转化相关基因
由表3可以看出,不同退化程度草甸的土壤氮素转化微生物基因丰度差异显著。其中,nifH基因的丰度变幅最大(每g干土6.88×103~2.76×109个拷贝数),且随退化程度的加剧而逐渐下降,与未退化草甸相比,重度退化草甸的nifH基因丰度降低了78.1%。amoA-AOA和amoA-AOB基因丰度分别为5.24×103~3.06×109和7.02×103~2.96×109个拷贝数,两者变化趋势相同,均随退化程度显著降低(p<0.05),中度退化和重度退化草甸的土壤amoA-AOA基因丰度无明显差异(p>0.05),但均显著低于未退化和轻度退化草甸。nirK和nirS基因变化趋势与nifH相反,随退化程度的加剧而增加,重度退化草甸的nirK和nirS丰度最高,显著高于未退化草甸(p<0.05)。轻度退化和中度退化草甸土壤中narG基因丰度与未退化草甸无显著差异,但显著低于重度退化草甸(p<0.05)。不同退化草甸之间nosZ基因丰度无显著差异(p>0.05)。
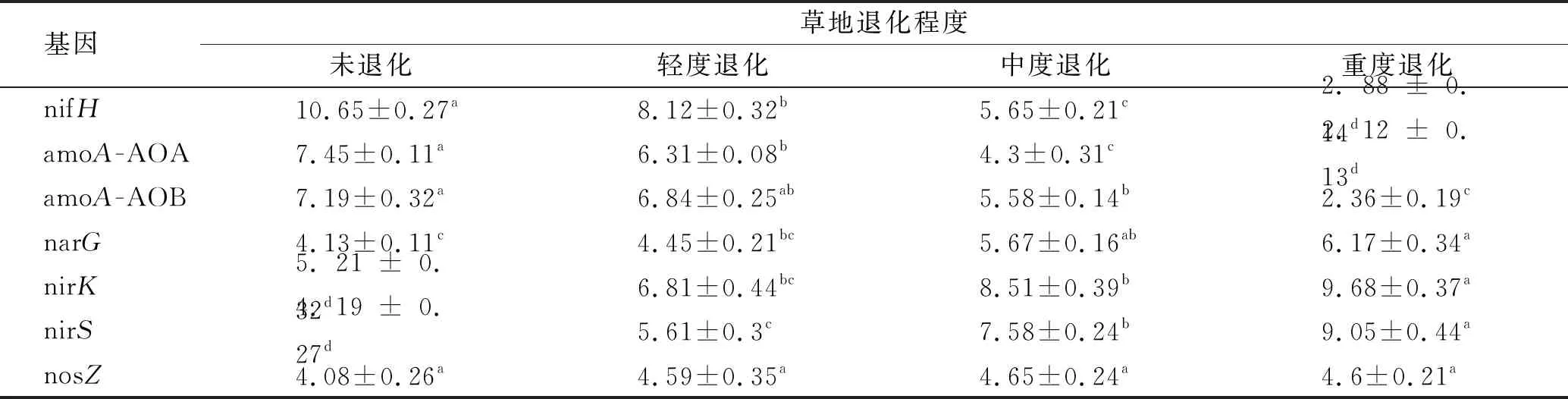
表3 不同退化草地土壤氮转化微生物功能基因丰度
2.3 氮素转化微生物基因与土壤理化性质的相关性
相关性分析表明(见表4),nifH基因与土壤pH值呈极显著负相关(p<0.01),与水分、有机碳、硝态氮、铵态氮及速效钾呈显著正相关(p<0.05)。
由表4可知,amoA-AOA和amoA-AOB基因丰度与土壤含水量的相关性最大,相关系数分别为0.76和0.81。nirK和nirS基因与pH值呈显著正相关(p<0.05),而与土壤含水量、有机碳、硝态氮、铵态氮和全磷呈显著负相关。narG则与铵态氮呈显著正相关。
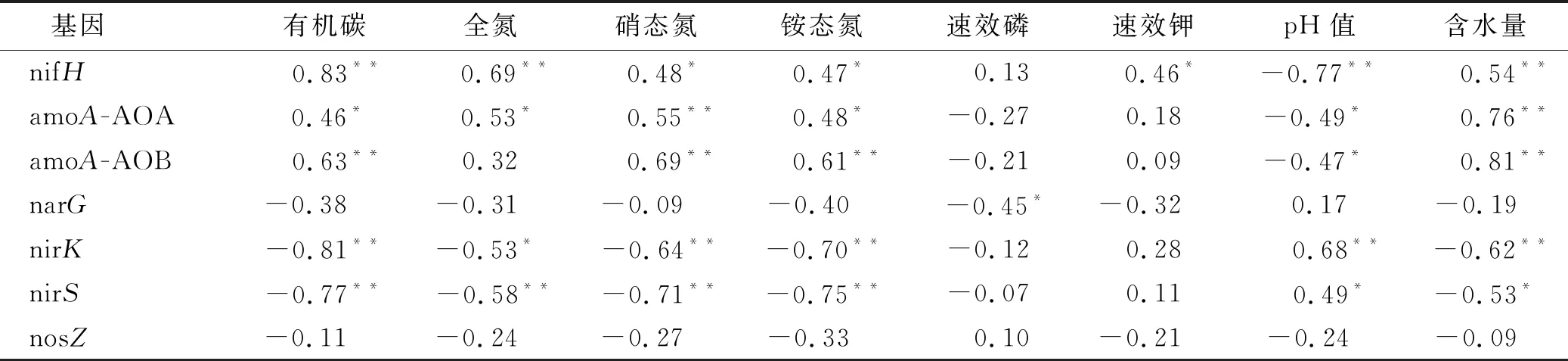
表4 土壤理化性质与氮素转化微生物基因丰度的相关性
RDA分析表明(见图1),土壤理化特性解释了53.2%氮转化功能基因丰度变异,其中第1轴解释率为52.7%,第2轴解释率为10.5%。土壤有机碳,pH值和含水量是影响高寒草甸土壤氮素微生物基因的主要因素。其中,有机碳和pH对nifH基因变化的影响较大,含水量对amoA-AOA和amoA-AOB基因影响较大,pH值对nirK和nirS基因影响较大。
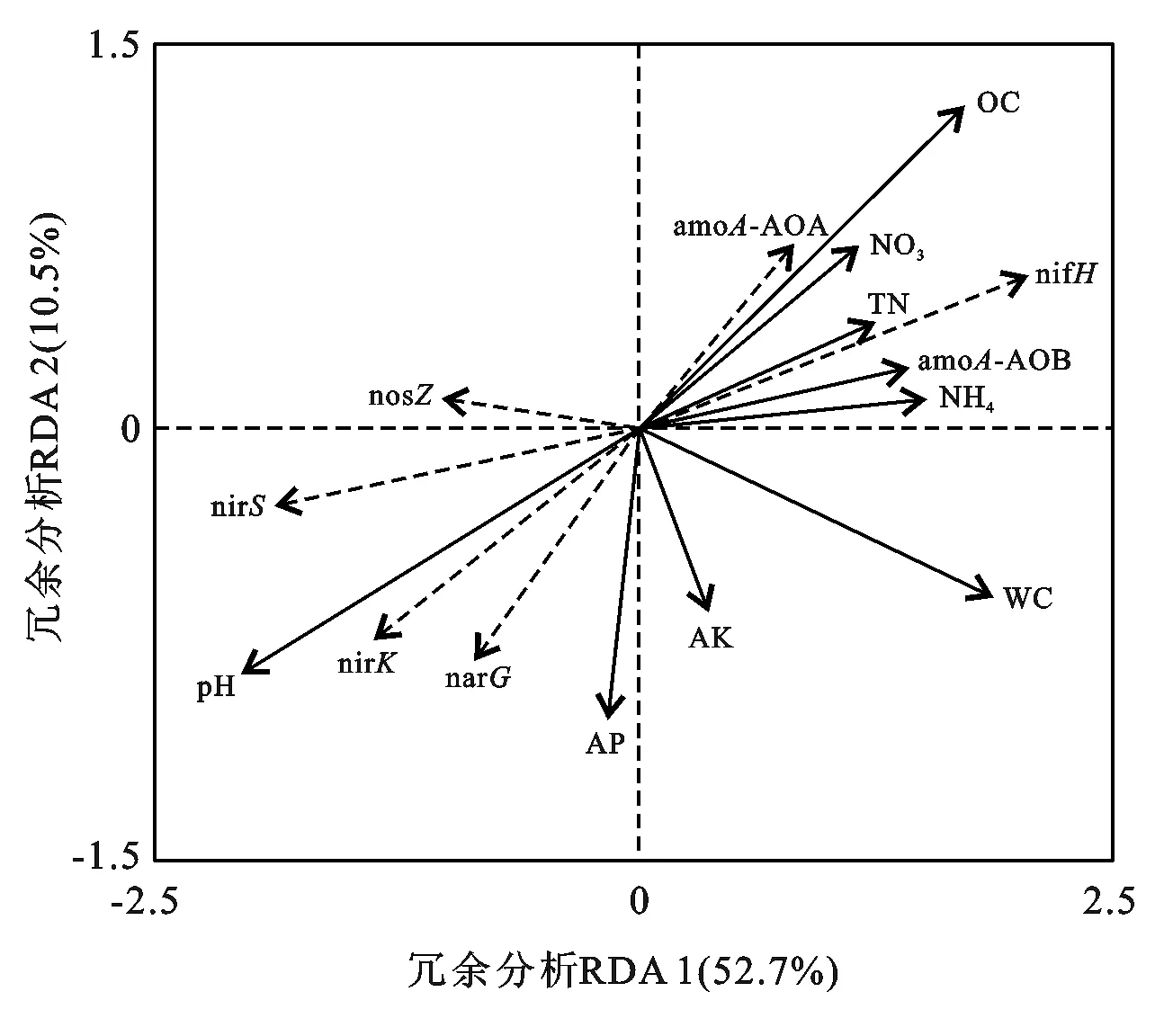
注:OC为有机碳; TN为全氮;为铵态氮;为硝态氮; AP为速效磷; AK为速效钾; WC为含水量; 虚线箭头线为微生物基因丰度; 实线箭头线为土壤理化特征。
3 讨 论
在土壤—植被体系中,土壤制约植被,尤其是土壤养分会引起植物群落结构及物种多样性的改变[14]。近年来,由于超载放牧等人为干扰及气候变化异常等多重因素的影响,草地沙化、黑土滩化、植被稀疏化等植被和土壤退化过程不断加剧,高寒草地生态功能严重受损。本研究表明,随着退化程度的加剧,土壤有机碳、全氮、铵态氮和硝态氮含量均显著降低,这与王玉琴等[15]、郭小伟等[16]等研究结果一致,其原因与地上生物量的减少密切相关。高寒草甸退化导致生物量减少,枯落物也相应减少,导致输入到土壤中的有机质下降,土壤中的碳氮主要来源于有机质,因此土壤中的碳氮含量显著降低。前人研究表明,植被覆盖度与土壤含水量呈显著负相关关系[17-18]。植物盖度降低必然导致土壤裸露面积增大,引起地表蒸发量增大,进而导致含水量降低以及土壤pH值增加,因此在本研究中,土壤含水量随退化程度加剧显著降低,而pH值则显著增加,这与高海宁等[19]研究结果一致。王玉琴等[15]发现,高寒草甸在退化过程中,土壤铵态氮和硝态氮的含量逐渐增加,推测其原因是啮齿动物排泄物的增加,导致速效氮含量增加。然而,我们发现土壤铵态氮和硝态氮随草甸退化程度的加剧而显著降低,与王玉琴等[15]结果相反。由于氮素的转化易受环境条件的影响,因此这种结果差异可能与不同草地类型以及人为或动物的干扰程度有关。磷、钾是植物生长所必需的营养元素,在植物生命活动中发挥着重要作用[20]。本研究中,随着退化程度的加剧,土壤速效磷、速效钾均呈降低趋势。造成这一现象的原因是退化草地中土壤微生物减少,使得植物残体的分解速率降低,导致土壤中速效磷和速效钾含量降低。
4 结 论
青藏高原高寒草甸退化导致土壤有机碳、全氮、硝态氮、铵态氮及水分含量降低及pH值升高。高寒草甸的退化降低了固氮nifH基因、氨氧化amoA-AOA和amoA-AOB基因丰度,但增加了反硝化narG,nirS和nirK基因丰度。土壤有机碳,pH值及水分是影响高寒草甸土壤氮素转化微生物基因的主要因素。
