云南省热带、亚热带和亚高山森林凋落物层中弹尾类群落组成的研究
2020-08-03徐艺逸
徐艺逸
(1.中国科学院西双版纳热带植物园热带森林生态学重点实验室,云南 勐腊 666303;2.中国科学院大学,北京 100049)
1 引言
土壤动物作为重要分解者,在生态系统的物质循环与营养供应中起着重要的作用。一方面土壤动物积极同化各种有用物质以建造其自身,另一方面又将其排泄产物归还到环境中不断改造环境。在土壤食物网中,土壤动物主要通过取食作用影响其他生物的数量以及群落结构。土壤食物网在有机质分解、碳存贮、土壤结构调节、病虫害控制以及矿物质养分循环中起着重要作用[1]。弹尾类在土壤中十分常见,与螨类和线虫同属于3种最常见的优势土壤动物类群。弹尾类对环境的变化十分敏感,土壤温度、土壤pH值和土壤有机质含量等都可对其群落组成产生影响,所以弹尾类也可作为环境指示生物[2~5]。
关于土壤动物群落组成的研究多关注不同海拔的区别,其结果主要分为两类:①对乌鲁木齐雅玛里克山(温带)土壤动物群落的研究表明:随海拔上升,土壤动物数量、多样性呈现递减趋势,不同海拔土壤动物群落间共有度较高,相似性差异不大,也有研究显示热带弹尾类多度和亚热带东部山地大、中小型土壤动物(土壤弹尾虫与土壤螨类)多度也有相同的结果[6~8];②衡山森林生态系统(亚热带)土壤动物密度最大出现在中间海拔段,海拔的升高或者降低土壤动物相应减少,在大兴安岭森林(寒温带)中小型土壤动物种类、数量和黄山(亚热带)土壤弹尾类的数量均有相似结果[9~11]。
虽然关于弹尾类群落组成在不同海拔的研究较多,但很少有人进行跨气候带与海拔的交叉研究。云南省分布着热带(西双版纳补蚌)、亚热带(哀牢山镇沅千家寨地区)和亚高山(丽江玉龙雪山)3种典型气候带,它们处于同一经度(E 101°)的不同纬度上,这很好的排除了因经度差异造成的一系列影响。因气候带的不同,导致了土壤温度、土壤pH值和土壤有机质含量等差异将影响弹尾类的群落组成情况。本研究意在回答如下问题:不同气候带弹尾类的群落组成情况;影响不同气候带弹尾类群落组成的环境因子是否相同。
2 研究地区与方法
2.1 样地概况与设置
本研究样地设置于西双版纳补蚌(热带)、哀牢山镇沅千家寨(亚热带)和丽江玉龙雪山(亚高山带)。
西双版纳傣族自治州位于云南省西南部(21°08′~22°36′N,99°56′~101°50′E)。海拔范围从澜沧江河谷的最低点491 m至最高峰2429 m。年平均气温为21.8 ℃,年平均降雨量为1493 mm[12],每年5月到10月为雨季,11月到次年4月为旱季。海拔1000 m以下的河谷地区为砖红壤。主要的森林类型为热带季节雨林、热带季雨林和热带山地常绿阔叶林等。原始林的林冠高达50~60 m,由望天树(Parashoreachinensis)占优势,中下层乔木主要有毛猴欢喜(Sloaneatomentosa)、绒毛番龙眼(Pometiapinnata)、假海桐(Pittosporopsiskerrii)、云树(Garciniacowa)、蚁花(Mezzettiopsiscreaghii)、染木(Saprosmaternata)等,次生林主要由壳斗科(Fagaceae)植物组成[13]。
哀牢山国家级自然保护区(镇沅片区)位于云南省中部哀牢山地区(24°7′N,101°14′E),海拔2147 m,相对高度在2700 m以上。呈现雨季(5~10月)和旱季(11月至翌年4月)的交替。该地年平均降雨量1931.1 mm,相对湿度86%。年平均气温11 ℃,最冷月平均气温(1月)4.8 ℃,最热月平均气温(7月)16.5 ℃,低温极值-8.0 ℃,高温极值24 ℃[14,15]。海拔2000~2700 m的植被类型主要为中山湿性常绿阔叶林,优势植物包括壳斗科(Fagaceae)、木兰科(Magnoliaceae)、樟科(Lauraceae)、山茶科(Theaceae)等[16]。
玉龙雪山位于滇西北丽江县境内(27°10′~27°40′N,100°10′~100°20′E)。最高峰扇子陡海拔5596 m,相对高差3846 m。该区域年降水量935 mm,6~10月为雨季,这5个月的降水量为年降雨量的90%以上,11月至翌年5月为干季。年平均温度12.79 ℃,最热月为7月,月均温17.9 ℃,最冷月是1月,月均温5.9 ℃[17]。森林由下而上分别为云南松林、丽江云杉林、大果红杉林和冷杉林[18,19]。从低海拔至高海拔,土壤类型依次为棕壤(2800~3200 m)、棕色针叶林土(3200~3600 m)、高山草甸土(3600~4200 m)[20]。
样地设置:热带、亚热带和亚高山气候带间高差为600 m,在每个气候带内设置4个海拔样带,海拔样带间的垂直高差200 m,其中,热带海拔梯度为800 m、1000 m、1200 m、1400 m,亚热带海拔梯度设置为2000 m、2200 m、2400 m、2600 m,亚高山海拔梯度设置为3200 m、3400 m、3600 m、3800 m。每个海拔带上设置4个20 m × 20 m的样地。
2.2 土壤动物采集与鉴定
在样地内随机选择3个0.6 m × 0.6 m的小样方抓取其中的全部凋落物于布袋中,做好标签,三处合并为一个混合样。将野外采集的凋落物样品置于烘虫装置中,利用土壤动物惧怕高温和光亮的特性将其烘烤出来,落入酒精中即得到土壤动物的酒精浸泡样本。依据《中国土壤动物检索图鉴》《中国亚热带土壤动物》《中国土壤动物》等资料进行弹尾类的分类鉴定(属)并计数[21~23]。
2.3 环境数据采集
土壤温度:2017年5~10月在样地内布置的土壤温度自动记录设备(iButton纽扣式温度计,每个小时自动记录1个土壤温度数据,10 cm深),获得每个样地的雨季土壤平均温度(T-rain)、雨季土壤极端高温(T-maxR)与雨季土壤极端低温(T-minR)。
土壤水分:在取土壤动物的同时,随机选择3个小样方,分别用环刀取100 cm3的土壤放入铝盒,使用烘箱烘干法(105 ℃)测定土壤含水量(soil moisture)。
土壤pH值:使用pH测量仪测定。仪器型号为:雷磁PHS-25数显型pH计。
土壤容重(Bulk density):容重法[24]。
土壤密度:密度计法。仪器型号为:TM-85密度计。
土壤孔隙度(Soil porosity):通过容重和比重计算获得[24]:

凋落物厚度(Litterfall amount):用钢尺直接测量,调查记录野外获取凋落物层土壤动物的3个样方的凋落物平均厚度。
2.4 数据处理
考虑到生物群落组成多为非线性,对不同海拔梯度上的弹尾类群落组成的间接排序采用NMDS方法,直接排序采用CCA方法。CCA分析采用前置逐步回归法筛选具有显著相关的环境因子,通过模拟999次检验排序轴的显著性。
所有分析在R软件(Version 3.3.2)中进行,主要用到的R包有“vegan”,“ade4”,“packfor”等。
3 结果与分析
3.1 不同气候带弹尾类组成概况
热带共采集弹尾类2431只,隶属于10科27属。优势属有4个,其中球角虫兆属(22.1%)数量最多,其余优势属为近缺虫兆属(21.0%,等节虫兆科)、刺齿虫兆属(15.9%,长角虫兆科)和拟缺虫兆属(14.2%,等节虫兆科),这4个属合计占热带弹尾类总体捕获量的73.2%;亚热带共采集弹尾类2315只,隶属于8科21属。优势属有2个,其中符虫兆属为绝对优势属(40.1%),另一个优势属为近缺虫兆属(13.1%),共占亚热带弹尾类总体捕获量的53.2%;亚高山共采集弹尾类14404只,优势属有两个,其中绝对优势属为符虫兆属(46.5%),另一优势属为棘虫兆属(28.3%,棘虫兆科),这两属占亚高山弹尾类总体捕获量的74.8%。
3.2 弹尾类群落的间接排序(NMDS)分析
NMDS分析(图1)表明,在3个气候带上,弹尾类的物种组成在NMDS1轴能较好的区分,热带和亚高山气候带的物种组成有明显差异,亚热带作为过渡区域与热带和亚高山物种组成均有部分重叠。在气候带内部不同海拔段弹尾类的物种组成差异不明显。

图1 弹尾类物种组成在不同气候带的差异,基于Jaccard相似性指数(a)和Bray-Curtis相似性指数(b)的非参数排序(NMDS)结果
3.3 弹尾类群落的直接排序(CCA)分析
对全部海拔段的弹尾类群落组成与环境因子进行CCA分析(图2)发现,CCA1轴和CCA2轴共同解释了33.91%的物种信息量,显著相关的环境因子为雨季土壤平均温度、雨季土壤极高温度、土壤pH值和凋落物厚度。凋落物凋落物厚度和土壤pH值与CCA1轴呈正相关,而雨季土壤最高温和雨季土壤平均温度和CCA1轴呈负相关。CCA1轴的正负轴较好的分割了高海拔和低海拔区域,高海拔(≥3200 m)大致分布在CCA1正轴,低海拔(≤2200 m)分布于负轴。凋落物厚度明显与CCA2轴呈正相关,土壤pH值则呈负相关。在海拔1000 m、1200 m、1400 m、2000 m处弹尾类群落组成主要受雨季土壤极高温度和雨季土壤平均温度影响。在海拔3600 m、3800 m处弹尾类群落组成主要受高土壤酸碱度和凋落物厚度影响。

图2 整体海拔弹尾类群落组成与环境因子的CCA排序
对热带地区不同海拔的群落组成与环境因子进行CCA分析(图3)发现,CCA1轴和CCA2轴共同解释了27.97%的物种信息量。显著相关的环境因子为土壤pH值、凋落物厚度和雨季土壤极低温度。土壤pH值和雨季土壤极低温度与CCA1轴呈正相关,而凋落物厚度和CCA1轴呈负相关。CCA1轴的正负轴较好的分割了高海拔和低海拔区域,低海拔(800 m)大致分布在CCA1正轴,高海拔(1000 m、1200 m、1400 m)分布于负轴。凋落物厚度明显与CCA2轴呈正相关。在海拔1000 m处弹尾类群落组成主要受凋落物厚度影响。在海拔3800 m处弹尾类群落组成主要受土壤酸碱度影响。

图3 热带不同海拔弹尾类群落组成与环境因子的CCA排序
对亚热带地区不同海拔的群落组成与环境因子进行CCA分析发现(图4),CCA1轴和CCA2轴共同解释了33.28%的物种信息量。显著相关的环境因子为雨季土壤平均温度和雨季土壤极高温度。雨季土壤极高温度与CCA1轴呈正相关,而雨季土壤平均温度和CCA1轴呈负相关。雨季土壤极高温度和雨季土壤平均温度与CCA2轴呈正相关。在海拔2200 m处弹尾类群落组成主要受雨季土壤极高温度和雨季土壤平均温度影响。
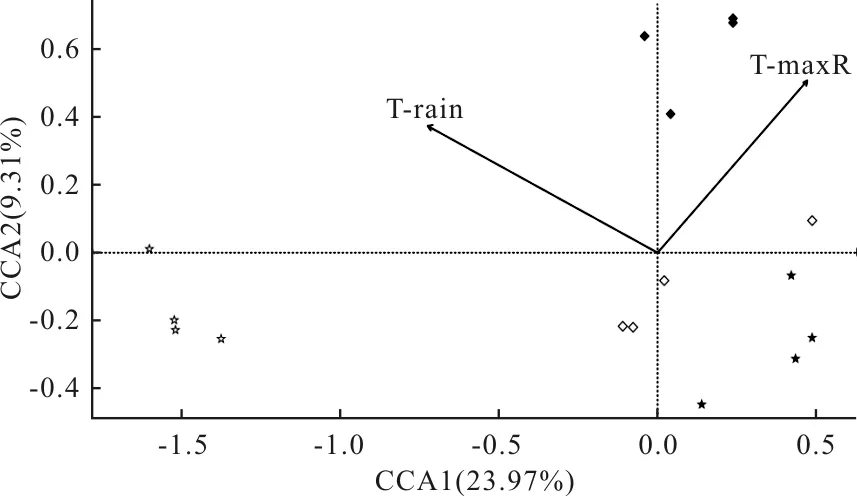
图4 亚热带不同海拔弹尾类群落组成与环境因子的CCA排序
对亚高山气候地区不同海拔的群落组成与环境因子进行CCA分析发现(图5),CCA1轴和CCA2轴共同解释了27.97%的物种信息量。显著相关的环境因子为凋落物的量、土壤孔隙度、土壤pH值。土壤pH值与CCA1轴呈正相关,而凋落物厚度和CCA1轴呈负相关。凋落物厚度、土壤孔隙度和土壤pH值明显与CCA2轴呈负相关。在海拔3800 m处弹尾类群落组成主要受凋落物厚度、土壤孔隙度影响。

图5 亚高山不同海拔弹尾类群落组成与环境因子的CCA排序
4 讨论
从NMDS分析图可以看出,不同气候带弹尾类的群落组成有所不同,热带和亚高山的物种组成有明显差异,亚热带作为过渡区域与热带和亚高山物种组成均有部分重叠。这可能是由于亚热带本就属于热带和亚热带的过度气候类型,气候特点处于二者之间,弹尾类受环境的影响,其群落组成与其他两个气候带均有相似部分。在气候带内部不同海拔段弹尾类的物种组成差异不明显,这可能是由于取样处于雨季末期,温度和土壤含水量等条件都较适宜弹尾类的生长,环境胁迫较少。
由整体弹尾类群落的直接排序(CCA)分析可以看出,亚高山弹尾类的群落组成的主要相关环境因子为土壤pH值和凋落物的厚度,可能由于亚高山气温常年低于其他两个气候带,而本研究取样于雨季,限制亚高山地区的弹尾类生长的温度胁迫消失,就体现出土壤pH值和凋落物的厚度(土壤有机质)对弹尾类的群落组成的影响,这与前人发现的对弹尾类群落组成的影响因素一致[3~5];热带弹尾类的群落组成的主要相关环境因子为土壤平均温度和土壤极高温度,可能是由于热带在3个气候带中年均温最高,而雨季的土壤温度在全年中又为最高,雨季平均温度和雨季极高温可能成为弹尾类的限制因子,之前的研究也发现了弹尾类的群落组成与温度密切相关[2,7];亚热带弹尾类的群落组成与本研究所涉及的环境因子相关性较小,可能由于其属于其他两个气候带的过渡气候类型,弹尾类的群落组成受到较复杂的环境因子影响,也可能本研究没有涉及到与其群落组成相关性较高的环境因子,有待进一步研究。分气候带的弹尾类群落的直接排序(CCA)分析可以看出,热带弹尾类群落组成主要与土壤温度、土壤pH值和凋落物厚度相关;在亚热带,与弹尾类群落组成相关的主要环境因子为土壤温度;而在亚高山,与弹尾类群落组成相关的环境因子主要是凋落物厚度、土壤孔隙度和土壤pH值。在气候带内部,与弹尾类群落组成的相关环境因子与整体排序的环境因子不同,可能是由于弹尾类生活在凋落物中,群落组成受小生境的影响更大。
5 结论
热带与亚高山弹尾类群落组成相差较大,亚热带作为过渡气候带,其群落组成与热带和亚高山均有相似部分;在气候带内部弹尾类群落组成相似度较高。从整体上看,土壤温度、凋落物厚度和土壤pH值是与弹尾类群落组成相关的主要环境因子;而不同气候带与弹尾类群落组成相关的环境因子有所不同,热带弹尾类群落组成主要与土壤温度、土壤pH值和凋落物厚度相关;在亚热带,与弹尾类群落组成相关的主要环境因子为土壤温度;而在亚高山,与弹尾类群落组成相关的环境因子主要是凋落物厚度、土壤孔隙度和土壤pH值。
