基于自由基对理论的仿生磁航向传感技术
2020-07-29程屹山刘文耀邢恩博
程屹山,刘文耀,邢恩博,唐 军,刘 俊
(1.中北大学电子测量技术国家重点实验室,太原 030051;2.中北大学仪器科学与动态测试教育部重点实验室,太原 030051)
0 引言
地磁场是指地球和周围空间之间存在的磁场,在地球近地空间内任意一点的地磁矢量都不同于其他地点,且与该地点的经纬度存在一一对应的关系。理论上只要确定该点的地磁场矢量即可实现全球定位,因此,地磁是导航领域的一种天然坐标系,如图1所示[1]。此外,地磁导航由于隐蔽性好、适应性强、误差不会随着时间产生累积等优势,能够在理论上实现全天候、全地域的导航服务。
然而,磁罗盘在多数情况下充当辅助角色,如何借助这一天然信息实现自主精确导航是目前研究的难题。在生物界,迁徙中的候鸟、海龟、鲑鱼可以依靠地磁导航,跨越上千千米准确归巢。正因为如此,生物磁导航机制一直吸引着相关领域的研究人员进行探究,而这一研究的突破也将成为仿生磁导航研究的关键性的一步。
本文将结合过去几十年来科学家对其研究的成果及进展进行评述,对动物感磁的自由基对机理(Radical Pair Mechanism,RPM)进行详细分析,并对与之相关的磁敏材料的制备及其磁场探测方式进行分析与总结。
1 生物磁响应机制
仿生地磁导航经过40余年的研究,有两种假说得到了学者们的广泛认可:磁受体假说和自由基对假说。前者以磁性颗粒为受体,后者以自由基对机理为核心。其中,自由基对机理是1978年由Schulten和Weller[2]教授将电子自旋理论引入到鸟类导航中提出的。这一假说认为电子自旋耦合和外磁场的共同作用导致单重态与三重态之间跃迁的方向不同,从而对化学反应的速率产生影响,使产率及产物的结构与磁场建立联系。
然而,自从这一机制被提出后就不断受到质疑,直到一种可以用作化学罗盘系统的蓝光感光蛋白被发现,才引起人们的广泛关注。研究发现,用于候鸟导航的自由基对对蓝光及近紫外光具有较强的光依赖性,其中Cry磁感蛋白很可能就是生物体内接受蓝光的磁受体。随后,Ritz等[3]在《Nature》上发表文章,通过低频振荡电磁波干扰实验证实了自由基对机理的正确性。之后,自由基对机理由Ritz和Wiltschkos等逐步完善,并得到了广泛的研究,被认为是最可能的、理论最完善的生物磁感应机理。
之后,一系列的实验和理论研究支持了基于隐花色素的自由基对假说:1)在鸟类视网膜细胞中发现了隐花色素,它能满足鸟类磁定向时的基本要求,同时,如果用于光信号处理的前脑区发生病变将会导致鸟类失去导航能力[4];2)鸟类利用地磁导航的能力与光的波长有关,主要以波长较短的蓝绿光为主[5];3)弱射频场可以破坏鸟的磁定向,并与自由基对作为研究对象的预测结果一致[6];4)利用时间分辨电子顺磁共振波谱观察到了隐花色素—光解酶中长寿命的顺磁性中间体,检测到自旋极化的瞬态自由基对信号,并根据瞬态自由基对的特征线宽、超精细分裂模式和共振磁场位置,确定了最终的电子供体是色氨酸残基(Tryptophan,Trp)[7];5)模型系统中的自由基对反应已经被证明能够对与地球一样弱的磁场做出响应[8-9]。
1.1 自由基对模型
随着研究者们对这一磁感机制的深入研究,其逐渐为人们所熟知:候鸟眼睛内存在光敏分子隐花色素,容易受光的激发接受一个质子,并从最邻近的色氨酸残基上夺得一个电子,形成[FADH·+Trp·]自由基对,进而演化产生自旋单重态和三重态。自由基对中未成对的电子自旋与核自旋相互作用且与外部磁场相互耦合,这些磁性相互作用使单重态的初始状态为非稳态。因此,自由基对电子自旋态在单重态(S)和三重态(T)之间表现出相干振荡,弱磁场也会影响其动力学。当外磁场介入后,磁场会影响单重态和三重态分子转换的速率,从而影响其产物及自由基对辐射退激发速率(寿命),这样就建立了弱磁和光信息转换的机理[4-5],被称为自由基对机理。另外,如果自由基对电子-核超精细耦合是各向异性的,那么外部磁场对其影响也是各向异性的,从而使单重态和三重态转换对外部磁场的方向敏感。
1.2 自由基对机理需满足的条件
研究发现,并不是所有能够形成自由基对中间体的化学反应都能够成为磁罗盘,而要满足以下5个基本条件:
1)反应中单重态和三重态中间体生成的化学产物不同,只有这样才能看出磁场对系间窜越的影响;
2)自由基的单电子必须与核子发生超精细耦合作用,因为超精细耦合不仅能让自由基对反应感受外磁场的影响,还能感受外磁场的方向;
3)作为磁罗盘的化学分子必须是具有空间取向的,因为处在溶液体系中的自由基对可以转动,从而抵消超精细耦合对外磁场的各向异性;
4)磁敏感的自由基对需由光激发形成;
5)自由基对中间体具有适当的量子寿命,寿命太长会被噪声干扰,而太短则不足以感受微弱地磁场的变化。
目前,自由基对机理成为了研究者们广泛接受的动物磁感受机制(图2)。近年来,生物学家们也发现了越来越多的诸如隐花色素之类的基于自由基对机制的磁敏感分子。因此,这些磁敏分子的磁敏性能引起了生物学家们的研究兴趣。

图2 候鸟的地磁导航机制Fig.2 Geomagnetic navigation mechanisms of migratory birds
2 基于自由基对机制的磁敏分子
2.1 蓝光受体—隐花色素
隐花色素作为生物体内最重要的蓝光受体之一,广泛存在于各种动植物体内,参与了多项生物体内生理功能的调节,与生物节律紧密相关。近年来发现,隐花色素是迁徙鸟类利用地磁导航的关键部件,也是最早发现存在于鸟类视网膜上且能够满足自由基对机理的磁敏蛋白分子。
在隐花色素的光化学反应过程中,黄素腺嘌呤二核苷酸(Flavin Adenine Dinucleotide,FAD)和3个色氨酸(TrpA、TrpB、TrpC)共同组成一个三联体分子,FAD受光照后处于激发态FAD*,电子经TrpC传递到FAD,形成自旋相关自由基对(Spin-Correlated Radical Pair,SCRP),如图3所示。研究发现,拟南芥隐花色素、鸟类隐花色素和海藻隐花色素等都可以形成长寿命的自由基对,并满足地磁条件下的磁场效应(Magnetic Field Effects, MFE)。

图3 天然隐花色素蛋白的结构Fig.3 Structure of natural cryptocyanin protein
越来越多的科学家们对各种隐花色素的磁场效应产生了极大的兴趣。先前,Marley等[10]研究了不同环境下,隐花色素的磁场依赖性对果蝇癫痫发作时间的影响,表明在蓝光(470nm)、磁场和Cry01/03作用下,果蝇癫痫的发作时间长。这是隐花色素的磁场效应对生物行为影响的一个强有力的证明。
那么隐花色素在不同溶液体系下的磁场效应会有怎样的变化?由于可能为磁感应分子的隐花色素不易被研究,而隐花色素光化学过程的关键组分是黄素,因此,通过研究黄素溶液体系的磁场效应能够对人们研究隐花色素的磁感应机理起到很大的借鉴作用。
目前,基于黄素的磁感蛋白模型体系被认为是最好的模型替代品,黄素是蓝光感光蛋白中重要的辅助因子。特别是其中的隐花色素,是动物磁感受自由基对假说的核心推定的磁感受器[11]。黄素容易与电子供体(例如色氨酸)发生依赖于pH值的光诱导电子转移,从而生成自旋相关的自由基对(Radical Pairs,RPs),这些自由基对可经历单重态和三重态之间的相干演化以及自旋选择性逆向电子传递,使得它们的光循环对外部磁场敏感。
由于在溶液环境中,适当的电子供体和受体可以发生光致电子转移形成SCRP,表现出磁场效应。之后,许多科学家对基于黄素溶液体系的磁场效应作了一系列研究。Tomoaki等[12]通过研究色氨酸和黄素单核苷酸(Flavin Mononucleotide,FMN)的混合溶液,发现其MFE仅为2%。随着Mohtat等研究提出了蛋白质与自由基之间的库伦作用对MFE有着重要影响,Tomoaki等[12]再次对黄素蛋白体系展开了更细致的研究。通过测量鸡蛋白溶菌酶(Hen Egg-White Lysozyme,HEWL)和FMN混合溶液体系,得到在水溶液中的MFE高达13%(250mT),而在加入NaCl溶液中的MFE降低为7%左右。
紧接着,生物学家研究发现,在胶团环境中的黄素溶液体系具有更好的环境相似性。于是Horiuchi等对此作了研究,发现在基于黄素溶液的胶束环境下,由核黄素(Riboflavin,RF)、FMN以及四丁酸核黄素(Riboflavin Tetrabutyrate,RFTB)和色氨酸形成的SCRP同样具有MFE,且在FMN胶团溶液中施加200mT磁场时的MFE约为3%~7%,RF/Trp体系可达15%(200mT),RF/Ind体系可达25%(200mT),而在高浓度环境下的RFTB/Ind体系可达40%(200mT)。
以上总结了基于黄素溶液体系的磁场效应的研究。为了更加全面地理解隐花色素磁感应机理,研究人员设计合成出各种类型的模型分子体系。由于隐花色素光化学过程的关键组分是黄素,因此隐花色素模型体系中应具有这个关键因素。
FAD是生物体系中重要的辅酶之一,并且自身就是一个由电子供体和电子受体组成的链连接系统,可在分子内发生电子转移(图4)。Lewis等[13-14]通过瞬态吸收方法测量了532nm处FAD的MFE衰减曲线的PH依赖性,当PH=2.3时,MFE峰值最高,且下降最缓慢;随着PH值的上升,MFE峰值逐渐减小,且下降越迅速;直至PH=8时,MFE接近于0,几乎呈一条直线。接着在532nm激光的激发下,测量了FAD的MFE在PH=2.3和PH=8时随外部施加磁场的变化,发现FAD的MFE在PH=8时随外部场增大上升迅速;在PH=2.3时上升缓慢,而这一变化正是由于FAD分子构象的变化导致的分子复合动力学的改变。

图4 FAD化学结构的表征Fig.4 Characterization of FAD chemical structure
FAD分子存在堆叠与伸展两种构象,核糖醇基-焦磷酸-核糖链间形成稳定的氢键网络,控制分子构象的转变[15]。FAD构象也受到溶液介电常数的影响,其荧光强度与介电常数呈线性关系,这一发现在生物研究方面具有重要价值。
通过对FAD的研究可以看出,溶液酸碱度以及介电常数的改变均能够影响分子动力学,从而导致MFE的改变。
实验已经证明在强磁条件下,隐花色素模型体系表现出了明显的磁场效应,但是如此高的磁场强度根本无法满足生物磁传感的实际应用。而在低磁场下,传统的单通吸收光谱法由于动态范围有限且通常需要测量大背景信号的微小变化而使其灵敏度受到限制。在实际生物系统中检测磁场效应时,这个问题变得尤为严重。
近年来,在基于宽带腔增强吸收光谱法方面,牛津大学的Hore等[16]开发出了一种新型的冷凝相BBCEAS MFE光谱仪(图5)。该系统能够在425~700nm整个可见光谱区间同时记录MFE,突破了较早的仅限于单波长测量的方法,并将其用于FMN和FAD两种隐花色素磁敏蛋白模型的研究。该仪器将来自超连续谱白光光源的光与由2个宽带涂覆的反射镜形成的简单光学腔耦合,以提供全可见光谱覆盖(425~750nm)。在近似十几mT的外部磁场下,测量基于黄素的磁感模型体系化学反应的磁场效应,可以得到最小可检测吸光度<10-7,从而允许检测到MFEs<10-4,并为实际检测生物样品中的MFE提供了可观的前景。

图5 用于磁场效应研究的BBCEAS系统Fig.5 BBCEAS system for the study of magnetic field effects
那么像地球一样大小的弱磁条件下,隐花色素蛋白是否也能表现出一定的MFE呢?
2014年,国防科技大学的杜现礼等[17]从昆虫细胞中分离纯化出了鸽子隐花色素蛋白,并发现在常温常压下该隐花色素蛋白能够对地磁大小的场强做出响应,而该技术与方法也是国内外的首例实现。通过对黑暗条件下解冻的C/CRY1蛋白样品进行瞬态荧光光谱的测试,发现当磁场为45μT时,瞬态荧光上升最快,衰减也最快;当磁场为55μT和75μT时,曲线右移;当磁场为142μT和285μT时,曲线左移。在排除了仪器本身受环境影响的情况下,表明磁场对蛋白分子FAD产生了影响。
鸽子隐花色素的成功提取以及对地磁信号的特异性响应为生物学的研究再一次迈出了关键性的一步。然而动物隐花色素的获取依旧较为困难,为了优化动物隐花色素的获取方法,2016年,同样在国防科技大学,袁晓霞等[18]对鸽子隐花色素(C/CRY1)系统的表达条件与纯化工艺进行了详细的分析、探索与优化,获得了大量纯化的C/CRY1蛋白,最终成功解决了鸽子隐花色素蛋白纯化效率低以及在获取时出现的辅基脱落的问题。
事实上,磁场对隐花色素中这种自由基对反应的影响已经在许多实验研究中得到证实。然而,这些研究几乎都使用了比地球多几个数量级的场,只有一项研究为低场效应提供了证明。Barton等通过实验证明了将DNA与一种修复紫外线诱发的病变的光解酶相结合能够对微弱磁场作出响应。但从中也发现了一些问题,例如,自由基对寿命极其短暂,且2个自由基的间距太小而导致极大的交换相互作用,这2个因素都不符合低场敏感的自由基对的要求;对地球强度场的各向异性的响应只记录在模型系统中。
2.2 富勒烯-卟啉-类胡萝卜素三联体(C-P-F)
为了使自由基对响应磁场的方向,必须固定至少一个自由基,以保留其各向异性的磁性相互作用。为了达到此目的,Meada等[19]选择了通过人工合成的由类胡萝卜素(C)、卟啉(P)和富勒烯(F)组成的三元组分子,结构如图6所示。经研究发现,该分子能够对近似地磁场(约50μT)的变化产生响应。
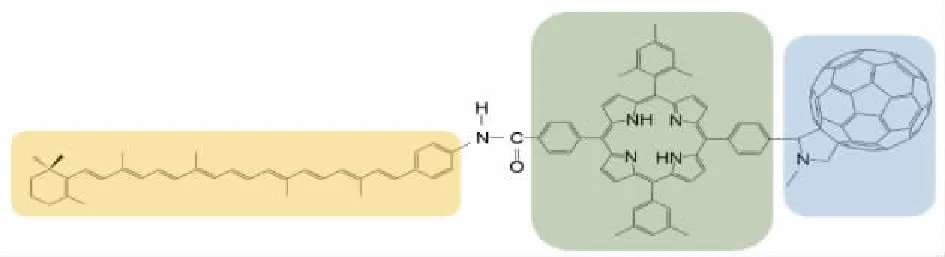
图6 C-P-F磁感应模型的化学分子结构Fig.6 Chemical molecular structure of C-P-F magnetic induction model
C-P-F分子最初是用来作为研究光合作用过程的有效的模型分子,虽然之前对黄素分子混合溶液体系的磁场效应研究不少,但是,还没有一种能够专门替代隐花色素的模型分子,而光合作用的模型替代分子经研究成为了隐花色素的一种很好的替代品。
C-P-F分子在绿光辐照下,内经电子转移,产生了自旋相关的单重态自由基对S[C*+-P-F*-]。然后,S[C*+-P-F*-]经历反向电子转移,直接转换为基态,其速率常数为ks,或者转换为激发的三重态,再退激发变为三重态自由基对,其速率常数为kT。最后一个过程是由2个未配对电子的磁相互作用控制的,是磁场敏感的步骤。正如对相关的三重态所观察到的那样,外加磁场通过改变自旋态的单重态特征来改变[C*+-P-F*-]的寿命,从而改变了kS和kT对整体动力学的相对贡献。
Meada等首先描述了外加磁场对各向同性溶液中自由基对消失动力学的影响,发现C-P-F的磁场效应的振幅随温度的升高而减小,且差分信号在200K以下呈双相分布。随后,测量了100ns与400ns处C-P-F分子的磁场效应随磁场强度的变化,发现当施加磁场强度大于1mT时,400ns处C-P-F的磁场效应表现出升高的趋势,而100ns处表现为下降的趋势,整体呈现出与预期相同的一种双相磁场效应。当施加类似于地磁场强度的场强时(B~50μT),显示出C-P-F吸收的变化高达1.5%。
要发挥化学罗盘的作用,自由基对磁受体必须对外部磁场作出各向异性反应。随后,Meada等使用对准样品证明了[C*+-P-F*-]的这种特性。图7所示为用于测量[C*+-P-F*-]磁场效应各向异性的实验装置示意图,磁场大小为3.1mT。测量结果发现,[C*+-P-F*-]的吸收显示出与θ明显的相关性,使用对准样品与光选择法测量的最大磁场效应分别为1.5%和5%。

图7 实验装置示意图[78]Fig.7 Schematic diagram of the experimental device[78]
C-P-F三联体的磁场效应各向异性的测量为生物磁传感的研究跨出了重要的一步。但是,进一步提高自由基对各向异性的磁灵敏度也是研究的难点所在。尽管研究者们对于优化其灵敏度的物理和化学性质尚不清楚,但单重态和三重态的重组率显然起着重要的作用。随后,Meada等[20]基于此特性对C-P-F分子的自旋选择动力学作了详细的研究,通过施加静态场、射频场和微波磁场对自由基对自旋态进行控制。最终发现,C-P-F分子的单重态重组率远大于三重态重组率(kS≪kT),显然这是C-P-F分子强磁效应的必要条件,但这是否也是对生物磁受体的要求还有待观察。
通过以上分析,C-P-F分子已被证明可用作化学罗盘,且反应产率能够表现出对磁场方向的敏感性。之后,为了进一步表征该系统,Meada等[21]再次开发出了一种时间分辨的低频反应检测磁共振(time-resolved Low-Frequency Reaction Yield Detected Magnetic Resonance, tr-LF-RYDMR)技术。实验中,通过改变所施加的静态磁场和36MHz振荡磁场的相对方向,发现静态磁场与振荡磁场的角度所引起的磁敏性极大程度上依赖于超精细耦合相对于静态磁场强度的大小。这一发现也为解释之前使用弱静态场(46μT)对动物进行研究的实验提供了一个至关重要的考虑因素。
以上实验基本表明,C-P-F三联体能够对地磁场产生响应,且对弱磁场也能够产生各向异性的响应。而为了进一步观察C-P-F对近似地磁场的各向异性的响应,Christian等[9]利用瞬态吸收光谱法,通过3对相互垂直的亥姆霍兹线圈产生的幅度恒定但方向变化的磁场,证明了C-P-F分子对微弱磁场(~1G)明显的取向依赖性。这也是C-P-F分子对近似地磁场各向异性响应的首例实现。
简而言之,C-P-F是目前通过人工合成的能够检测到地磁变化的磁敏蛋白。但由于实验不符合常温常压的条件,需在极低的温度下进行,因此,该磁感分子没有被广泛用于实际应用当中。
2.3 链状合成分子
探索能够代替生物体内磁感受器的模型体系的步伐并未停止。众所周知,电子转移对芘/N,N-二甲基苯胺(Py/DMA)的激基复合物荧光依赖于磁场。紧接着,许多科学家基于该自由基离子对作了研究,Roy等[22]通过研究不同溶剂中Py/DMA的MFE,发现不同的溶剂极性对其MFE影响很大,随着溶剂极性的不断增大,其MFE先增大后减小,在溶剂介电常数ε=15时,MFE达到极大值。Cohen使用Py/DMA的荧光执行一种新型成像,称为磁荧光成像(Magnetic Fluorescence Imaging,MFI)[23],可以通过强烈散射的介质(如磨砂玻璃)成像。然而,由于再生率和Py/DMA中自旋演变的微妙相互作用,MFE的大小对溶剂介电常数和极性非常敏感,这限制了MFI的效用。而通过链连接电子供体和电子受体的磁感分子能够增加重组的可能性以及MFE的大小,并在常温常压下表现出明显的磁场效应,尤其是当2个自由基的超精细耦合作用极不对称时,能够对外磁场做出各向异性的响应。为了便于合成,目前研究者们拟分别采用菲、芘、蒽等环状材料作为电子受体,二甲基苯胺(Dimethylaniline,DMA)作为电子供体,并通过控制链长而延长自由基对的退激发寿命达到μs量级,以便于能够感知磁场变化的信息。
早在20世纪80年代,Weller和Staerk以及他们的同事针对链状分子Py-(CH2)n-DMA展开了一系列磁场效应的研究,这些研究成果也促使相关领域的科学家们对此类磁感分子进行更深入的研究。随后,Yoshihisa等[24]于1996年合成出了共轭链连接的链状磁感分子Phen-(CH2)10-O-(CH2)2-DMA,并研究了该分子在不同环境下(链长、溶剂、温度)的磁场效应。研究发现,该磁感分子随着亚甲基数n(n=4,6,8,10,12)的不断增大,MFE(IB/I0)在B=0.35~0.59T时呈现不断增大的趋势;随着溶剂介电常数ε的增大(12<ε<37),MFE(IB/I0)在B=0.26T时呈现不断增大的趋势;在B>50mT时,随着温度的不断增加,n=7的链状分子的MFE(IB/I0)呈现不断减小的趋势,最后测量得出该分子在一定条件下的MFE高达140%。紧接着1997年,Yoshihisa等[25]通过合成不同链长的磁感分子,测量了强磁场对Phen-(CH2)n-O-(CH2)2-DMA(n=6~8,10,12)的荧光强度的影响。发现在施加磁场大于1T时,磁感分子的MFE会出现反转。基于对链状磁感分子在不同环境下的磁场效应的测量结果,越来越多的科学家们也开始对这种人工合成分子进行探索。
如何提高链状磁感分子的磁感灵敏度是生物磁传感技术发展的关键一步。蔡建明受超精细耦合对外磁场各向异性理论研究的启发,发现利用铁磁性纳米颗粒设计材料,能够改善各向异性的超精细耦合,提高磁感灵敏性能。Cohen等[26]根据理论研究也发现,适当设计的纳米球状梯度场,可以显著提高化学罗盘的性能。例如直径10nm的Fe3O4球在其表面产生约600G/nm的梯度场,该梯度场约为电子-核自旋耦合产生磁场的10倍,且远远超过用宏观线圈产生的效果。若梯度场在某一个自旋的位置处强,而在另一个自旋处近似为0,就可以定向地发生磁场耦合,消除小空间区域中纳米结构的场互扰,从而实现纯分子罗盘所需的强的各向异性超精细耦合,能够显著提高磁场感应灵敏度。因此,在设计真实器件时,可以通过在硬铁磁性纳米结构附近施加空间均匀的偏置场,而产生超精细耦合,代替局部强梯度磁场[17],实现使用自由基对反应映射纳米磁场。

表1 三种磁敏分子的性能及优缺点Tab.1 Performance, advantages and disadvantages of three kinds of magnetic sensitive molecules
基于磁性纳米颗粒的这一特性,Cohen等[27]仿照Yoshihisa等合成链状磁感分子的过程,同样地合成出了类似的Phen-(CH2)12-O-(CH2)2-DMA磁感分子,并通过在磁感分子附近设置铁磁性纳米结构,利用其自由基对反应机理绘制出了不同形状的铁磁纳米颗粒周围的磁场分布。以圆形铁磁颗粒为主,量化了从结构边缘扩展13μm的荧光角,结果显示,在0°和180°时MFE最大,且最大MFE达1.7%,在90°和-90°时MFE最小。
基于此结果,为了真正地实现三维轮廓分析,Cohen等[28]引入了一种利用双光子共焦荧光来实现荧光成像的方案。研究发现,利用双光子激发Phen-(CH2)12-O-(CH2)2-DMA同样能够保持磁场效应,并利用此属性绘制了三维铁丝周围的磁场分布。以上实验结果基本表明,基于电子受体菲和电子供体DMA连接的磁感分子能够在常温常压下表现出良好的磁场效应以及对磁场各向异性的响应,而通过改变电子受体或电子供体是否能够进一步增强磁感分子对磁场的响应呢?Kattnig等[29]通过研究链状供体-受体系统的自由基对动力学发现,MAnt-(CH2)16-O-(CH2)2-DMA分子在B=75mT时,MFE达37.5%。Lee等[30]通过对合成出的新型磁感分子Py-(CH2)12-O-(CH2)2-DMA的研究发现,应用广泛使用的355nm的紫外光激发时,该分子显示出比Phen-(CH2)12-O-(CH2)2-DMA分子更明显的MFE,并且所需的检测手段更为简便,所得的信噪比(Signal/Noise,S/N)更高(图8)。同时这一实验结果也能启发相关领域的科学家们探索更多类似的链状磁感分子,以求获得更高的磁场灵敏度。

图8 Py-(CH2)12-O-(CH2)2-DMA的化学分子结构Fig.8 Chemical molecular structures of Py-(CH2)12-O- (CH2)2-DMA
3 结论与展望
近年来,随着地磁测量技术的不断完善和发展,国内外越来越多的科学家致力于地磁导航的研究,并取得了丰硕的成果。但基于生物磁感应的仿生磁传感技术的研究仍未得到重大突破,而是否能够发现一种在常温常压下既能响应地磁大小又能响应地磁方向的人工磁感分子模型,是这一技术能否进一步得到跨越的关键。而通过将目前已合成出的链状分子与牛津大学等学者开发出的宽带腔增强法相结合,理论上能够实现常温常压下对微弱磁场变化的响应,但能否将其实现,还需通过实验进行研究。自由基对机制真的就是生物进行磁导航的内在机制吗?这一结论还需更多的实验依据加以证实。或许,当生物磁感应的内在机制得到了全面理解,仿生磁导航技术的发展将真正走向成熟。
仿生磁传感技术的研究一直是近些年国内外研究的热门领域,虽然眼前还有许多问题等待突破,但相信通过科学家们不断的努力与尝试,结合多学科交叉的技术,以及发展设计出更优秀的实验平台和模型体系,这些问题必将得到解决,将全面推动仿生磁传感技术发展并最终走向实际应用。
