Functional interactions among phytophagous insects, plants and microbial symbionts
2020-07-24WaleedAfzalNAVEEDJunaidAliSIDDIQUIYELongqinHUANGXiaolei
Waleed Afzal NAVEED, Junaid Ali SIDDIQUI, YE Longqin, HUANG Xiaolei
(State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China)
Abstract: Phytophagous insects and plants have intimate relationships with microbes that might influence their own ecological functions and interactions among them. Due to the advances in molecular studies and techniques, high diversity of heritable symbionts(e.g. bacteria, fungi) have been identified in both phytophagous insects and plants. Symbiotic microbes are present in hosts as a hologenome and influence biological processes such as nutrition, reproduction, defense and detoxification of toxins. Symbiotic relations may be competitive, mutualistic or parasitic. These interactions are frequently facilitated by other environmental microbes. Phytophagous insects can harbor symbionts in their bacteriocytes, gut, haemolymph and salivary glands. New molecular techniques and high throughput sequencing have increased the possibilities to explain the interactions even in low prevalence of symbionts. By summarizing recent evidence on different aspects such as the diversity of symbionts in phytophagous insects and plants and the interactions between them, mechanisms of insect-plant interactions, and detoxification of plant toxins by insect symbionts, here we emphasize the importance of understanding functional interaction networks among microbial symbionts, phytophagous insects and plants with a systems perspective, and provid our views on future research topics.
Key words: symbionts; hologenome; species interaction; mutualism; ecological function
1 Introduction
Insects and plants make diverse kinds of relationships with association of microbes that can affect the output of their own interactions(Sugioetal,2014;Gurungetal,2019). Plants are important mediators of interactions between their related microbes and insect communities(Van der Puttenetal,2001;Ohgushi,2005). Changes in plants incurred by one species have cascading effects on the interactions with other species(Figure 1C), influencing their abundances and the community structures(Ohgushi,2008). Different type of strategies has been evolved by insects in order to feed on plants by using mutualistic symbionts, that are important for direct and indirect relationships between herbivorous insects and their host plants(Figure 1A)(Barbosaetal,1991).
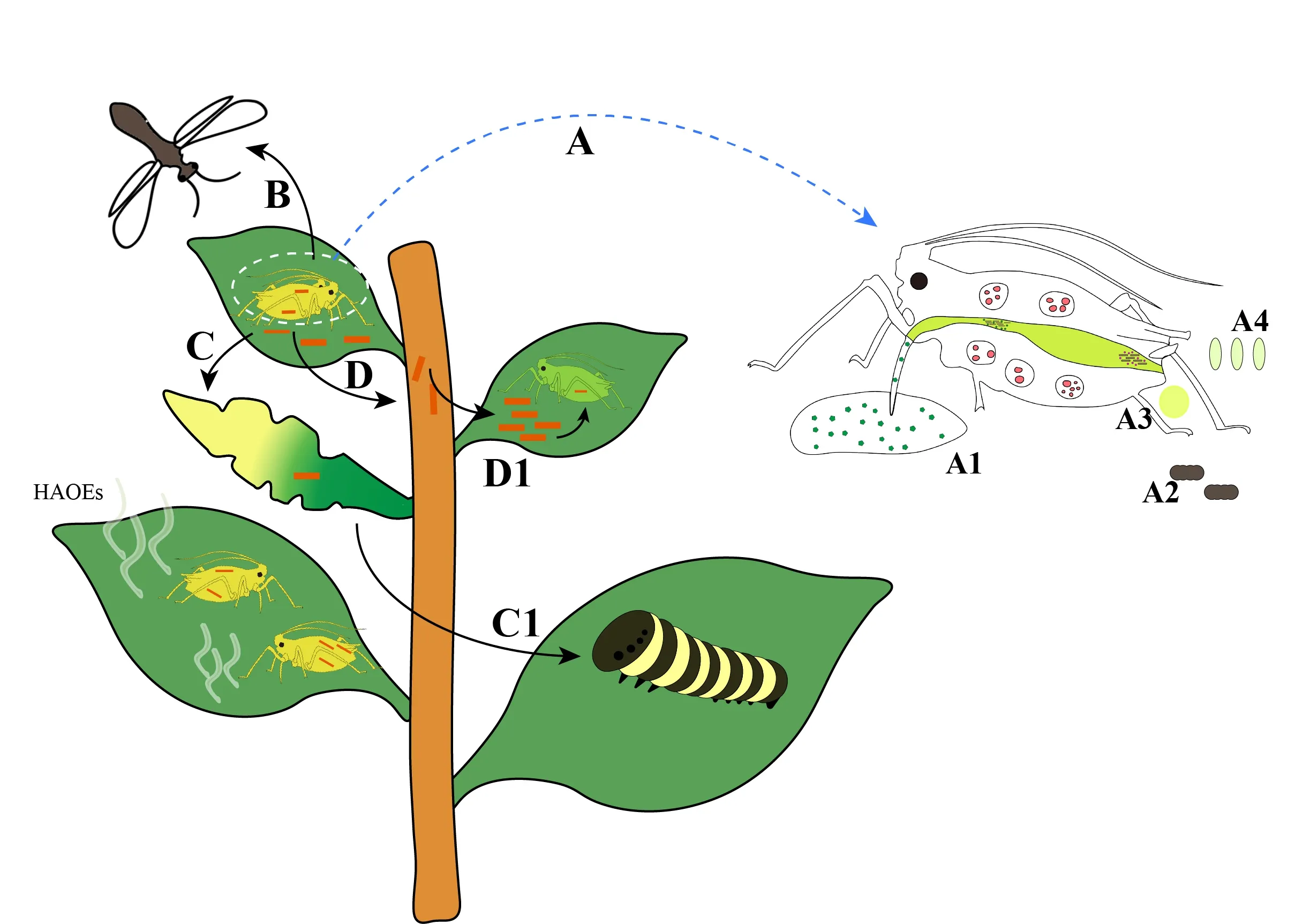
Note: Here we use aphid as model insect representing herbivorous insects. A. Different symbionts in insects differentially affect host plant use, and their locations within hosts(e.g. bacteriocytes, gut, oocytes) and transmission modes vary, for examples, in saliva(A1), on excretory material(A2), in honeydew secreted by insects(A3), and on eggs(A4). B. When the insects feed on the plants, symbiotic microorganisms affect the attractiveness of the host plants towards natural enemies. C. Symbiotic microorganisms can change the physiology of plants to avoid sharing of same host by other herbivores. D. During insect feeding on host plants, symbionts may be transmitted to the plants; and then the symbionts can be transmitted to symbiont-free insects by using the plant as an intermediary. Plants release several compounds like JA, CK, GA, ABA to suppress the insect feeding process, but herbivore associated organisms and elicitors(HAOEs) can interfere with these compounds and the plant defense system.
Symbiosis is a kind of ecological relationship between individuals of usually 2 different species that live together and form intimate associations(positive or negative). In the current review, we mainly focus on beneficial associations between microbial symbionts, phytophagous insects and plants. Symbiotic microbes are abundant in insect or plant hosts. They can form obligate associations in which both of symbionts depend entirely upon each other for survival or less intimate facultative associations. Microbial symbionts affect the development(Braendleetal,2003;Koropatnicketal,2004), nutrition(Liuetal,2014;Xieetal,2019), reproduction and speciation(Hurstetal,2000;Bandietal,2001), immunity(Macdonaldetal,2005), protection against natural enemies(Scarboroughetal,2005) and even species evolution(Gilbertetal,2012;Rosenbergetal,2016) of their hosts.
As a classic example, almost all aphids have an obligate bacterial symbiont(Buchnera) that can produce different essential amino acids and permits the insect to nourish on phloem(Jansonetal,2008;Gibsonetal,2010). The phloem-sap aphids(and other hemipteran insects) feeding on is deficient in essential amino acids and vitamins, that′s why they depend very much on obligate symbionts that supply essential nutrients. Fungi can also be nutritional symbionts and more probable to contribute sugars and sterols, comparing to the amino acids usually produced by bacterial symbionts. Yeast-like fungal symbionts of some hemipterans can produce enzymes and sterols for detoxification of allelochemicals and degradation of plant material(Jansonetal,2008;Gibsonetal,2010;Feldhaar,2011). Facultative symbionts can have prominent effects on host phenotypes(Oliveretal,2010;Feldhaar,2011). Infection by specific bacterium might be beneficial to the host under some conditions but harmful for other hosts or environments(Daleetal,2006).Hamiltonelladefensain the whiteflyBemisiatabaciis stated to proliferate the host′s fitness on low-nitrogenous foods(Suetal,2013). Some beetles attained plant cell wall lowering enzymes from bacteria or fungi. For example, b-fructofuranosidases(breaking down of the plant sucrose enzyme) were acquired by some coleopterans through horizontal transfer, possibly from the bacteria. The production of b-fructofuranosidases in insect′s cells(Keelingetal,2013;Pedezzietal,2014) allows them to feed more efficiently on plant sucrose. In obligate insect-microbe interactions, gene markers are usually used to investigate biogeographical patterns and evolutionary history of hosts(Liuetal,2013).
A three-way interaction between insect-plant and microbes has been highlighted in(Biereetal,2013). First is microbial intermediation between plant-insect interactions, such as change in morphology, phenology, and biochemistry of plants which affect the herbivore performance(Pinedaetal,2010). Second is insect intermediation between plant-microbe interactions, including accessibility of host plant tissues for symbionts(Stoutetal,2006) and the role of vectors of plant pathogens to introduce new diseases(Panetal,2012). Third is plant intermediation between insect-microbe interactions e.g. plant quality, nutrition for symbionts and herbivorous insect′s dependence on plants(Davisetal,2012). Some symbionts of insects are identified to affect relations between the natural and herbivores enemies. Symbionts can initiate volatiles that are used by some parasitoid species to locate the insect hosts(Dickeetal,1992;Godfray,1994;Dicke,1996). The interactions with natural enemies might be affected by the insect symbionts by their effect on the host′s resistance or susceptibility. The aphid endosymbiontRickettsiellawhich may affect the body color of aphids probably also influence higher trophic level interactions between aphids and natural enemies, for example, ladybirds and parasitic wasps(Tsuchidaetal,2010).
2 Insect as microbial community
Insects and a large group of microbes living within them form interesting multi-species communities. Microbes which live in insect body include bacteria, viruses, and fungi(Figure 1A). Recent studies have found that various non-pathogenic microorganisms can firmly inhabit hemolymph in a diversity of insects. Most known examples includeSpiroplasma, particularly inDrosophilaspecies, and secondary symbionts of the EnterobacteriaceaeHamiltonelladefensa, in aphids(Blowetal,2019). Symbiotic microbes may be different within different tissues of same organism, the diversity of microorganisms in holobionts depends on the habitat, immune system and different environmental conditions of different tissues(Figure 1A)(Rosenbergetal,2016). The stinkbugsMegacoptacribrariaandM.punctatissimagive a perfect example of insect symbiont participation as hologenome. Both species commonly found on legumes in Japan, but onlyM.punctatissimacan inhabit soybean and become pest of soybean. Experimental interchange of gut symbiontCandidatusIshikawaella capsulata between the two species indicated that this symbiont determined the pest status on soybean(Hosokawaetal,2007). The pea aphidAcyrthosiphonpisumis identified to be related with at least seven species of facultative symbionts and the chestnut weevilCurculiosikkimensiswith 6. Their frequency varied according to the insect feed on certain host plants(Oliveretal,2010;Tojuetal,2011;Zengetal,2019). Microorganisms able to complete their life cycles in insect hosts, fulfil their physiological needs and get nutrients and habitat from their insect hosts. For example, scale insects can be found in the middle side of the fungus which relate to several tunnels. FungusSeptobasidiumparasitizes scale insects by inserting their haustoria which is specialized feeding hyphae(Methéetal,2012). Hindgut is the region bearing largest microbial population. Ions and metabolites filtrate present in the hindgut play significant role in the microbial function and growth(Huangetal,2010). In latest research, insects also enable symbionts to be identified and used in latest pest management technologies. Many cuticles associated bacteria can proliferate and form their stable communities in insects e.g. cuticular invaginations of housing fungi mycangia required by insects stored and protected(Hulcretal,2012).
Insect-associated microbes adjust the plant defense strategies for the advantage of their hosts, and they may also act as elicitors to alter interactions between insect hosts and plants(Figure 1)(Wielkopolanetal,2016). Insects which feed on debris or wood are often detected with fungi with the ability to digest wood. For example, fungal symbionts in the bark beetles ofCurculionidaehave various benefits to their insect hosts(Douglas,2009), and lignin degradation might happen mainly due to fungal actions inAnoplophoraglabripennis(Cerambycidae)(Geibetal,2008).
Due to lack of some important nutrions such as essential amino acids and vitamins in related plant tissues, herbivorous insects have usually acquired nutritional symbionts that may stimulate host-plant specialization(Jansonetal,2008). Different hemipteran groups have unique bacterial symbionts, e.g. aphids withBuchnera, whiteflies withPortiera, psylids withCarsonella, and coccids withTremblaya(Hansenetal,2014). Some insects have two symbionts for nutritional provisioning, e.g. the conifer aphid genusCinarahaveBuchneraaphidicolaandSerratiasymbiotica, the mealybugPlanococcuscitrihaveMoranellaendobiaandTremblayaprinceps, and the leafhopperHomalodiscavitripennishaveBaumanniacicadellinicolaandSulciamuelleri(Table 1)(Tojuetal,2011). With the advances of molecular studies and techniques, more and more symbiotic microbes with important functions have been identified from insects and plants in recent years. We think it is important to summarize recent evidence on interactions among microbial symbionts, phytophagous insects and plants, which can connect the traditional disciplines such as entomology, insect and plant pathology, and microbiology, and improve our understanding of the evolution of biological complexity.
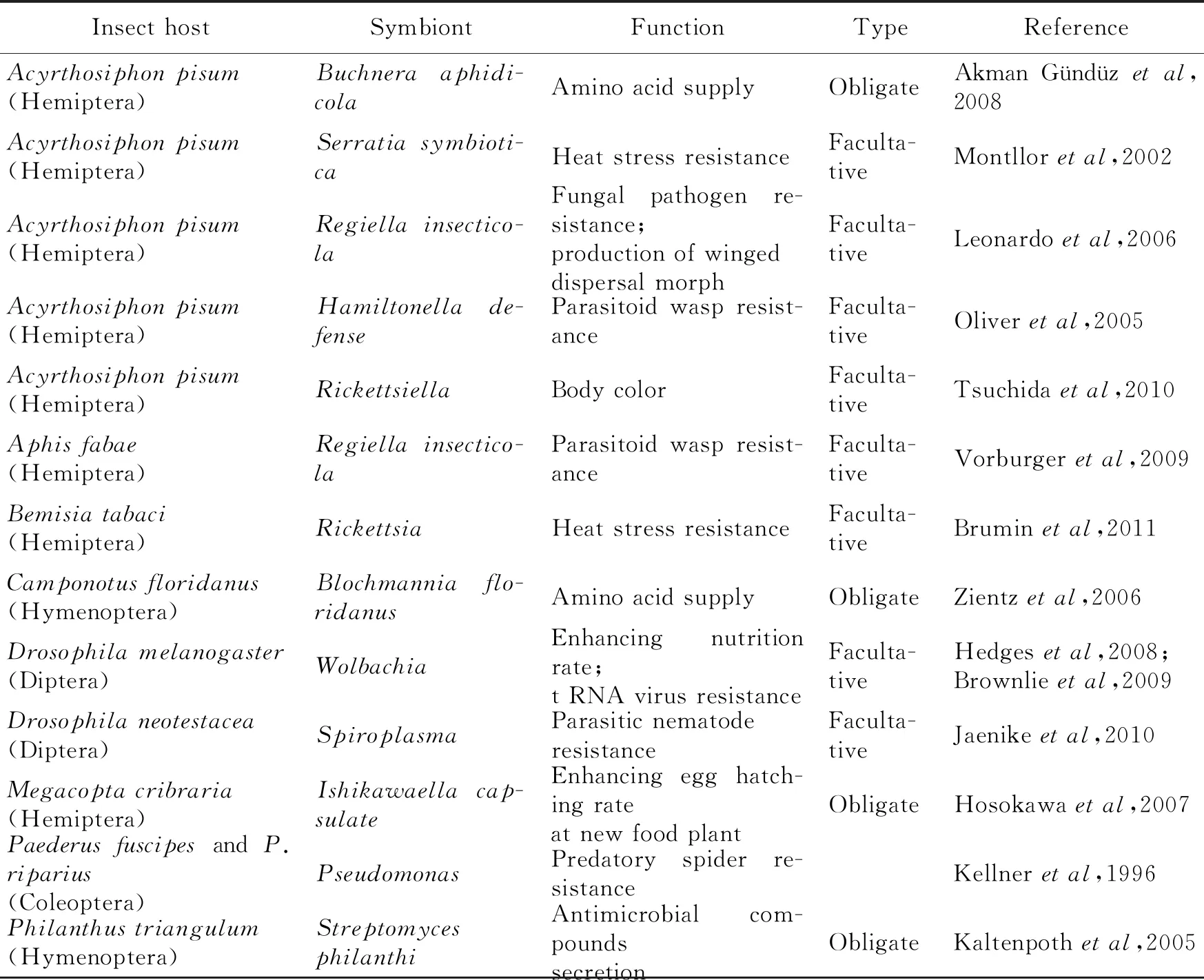
Table 1 Some important symbionts performing specific functions for insect hosts
3 Taxonomic diversity of insect symbionts
Herbivorous insects have a relatively narrow range of microbial communities, dominated by a few taxa(Colmanetal,2012;Jonesetal,2013). It was reported thatDrosophilaspecies harbor no more than 30 operational taxonomic units(OTUs), in contrast, mammalian guts have occupied by almost 1 000 taxa of symbionts(Chandleretal,2011). Previous studies showed that different insect orders may have a low bacterial diversity with 10-15 OTUs on average(Colmanetal,2012;Jonesetal,2013). Sap feeding insects like aphids, psyllids and whiteflies have been reported with 29-33 OTUs per sample, while xylophagous insects with highest gut bacterial groups almost 103 OTUs per sample, and leaf-feeders in-between with about 38 OTUs per sample(Russelletal,2013;Jingetal,2014). The bacteria present in holobiont in less than 4×107cannot be perceived by most existing methods, but such species cannot be discounted because they may be important for adaptation or evolution of the holobiont in any new environment(Rosenbergetal,2018).
Proteobacteria and Enterobacteriaceae are the most common microbes of herbivorous insects. Proteobacteria contains many obligatory as well as facultative insect symbionts such asBuchnerain aphids,Carsonellain psyllids andPortierain whiteflies(Moranetal,2008).LactobacillusandAcetobacterare common inDrosophila(Chandleretal,2011).FirmicutesandBacteroidetesare tightly linked with xylophagous insects, mainly those feeding on decayed wood(Colmanetal,2012).
Bacterial symbiont communities exist in different parts of host body. Many obligatory symbionts occupy specialized bacteriocytes, while facultative symbionts inhabit in different localities such as sheath cells and haemolymph. Bacterial communities can also inhabit in the salivary glands of their insect hosts(Bodyetal,2013). For instance,CandidatusLiberibacter, the causal organism ofCitrustrees, is established in salivary glands of psyllid vectors(Ammaretal,2011). This bacterium can also establish in salivary glands of mealybugs, but in which the strain is dissimilar from the one spread by psyllids and does not source of infection in host plants(Pitinoetal,2014).
4 Adaptation of insects to plants
The quality of host plant is important for the development and reproduction of herbivorous insects(Awmacketal,2002). Surface of plants have many structures like setae, hairs and thorns, which may cause restriction during the feeding(García-Munizetal,1998). And, some plants have toxic chemicals in their tissues. To overwhelm these problems, insects have adapted themselves to nourish on plants by the methods to sustain on plants and detoxifying the harmful compounds(Wielkopolanetal,2016).
Insects adapt to plants with or without the help of symbionts. The secretions of insects contain blend of components that help to feed on plants. Herbivorous insects have many organisms and promotors that work during feeding(Bonaventureetal,2011;Zhuetal,2014). Insect secretions are varied and can also contain enzymes b-glucosidase and glucose oxidase(Mattiaccietal,1995;Eichenseeretal,1999), peptides from consumed plant proteins(Schmelzetal,2006), cell-wall wastes pectins and oligo-galacturonides(Bergeyetal,1999), amended forms of lipids(Albornetal,2007;Hilkeretal,2010), or organism derived proteins and microorganisms(bacteria, viruses and fungi) that affect the outcome of plant-insect interaction. Some herbivorous insects may have effector elements that can conquer plant resistance reactions(Walling,2009). Mostly, these molecules destroy a jasmonic acid(JA) pathway, which is frequently activated in comeback to plant feeding insects(Chungetal,2013). These molecules may be existent in insect oral emissions or eggs(Consalesetal,2011;Atamianetal,2013).Leptinotarsadecemlineataharbors different bacterial symbionts in its secretion that can be delivered to plants. Flagellin resultant fromPseudomonassp. persuades salicylic acid(SA) pathway and overwhelm JA signaling pathway(crosstalk), accordingly decreases plant defense against insect(Figure 1)(Consalesetal,2011). Insects might also be linked with microorganisms that are harmful for plants. These plant pathogens may not only conquer plant reaction to the assistance of their insect host but can also modify plant physiology to fascinate vectoring insects to proliferate the chance of pathogen dispersion(Baietal,2009).
5 Transmission of insect symbionts mediated by plants
Insect symbionts are primarily transmitted vertically, but studies have also demonstrated that most facultative and even some obligate symbionts can be transferred horizontally(Jaenike,2012). The simplest way plant might be included as a passive surface, e.g., vertical transmission is not possible for some gut symbionts and positive inoculation of the following generation needs young ones to feed on contaminated material with the gut flora that is adult. InRiptortuspedestris, obligate gut symbionts are deposited in the capsules alongside the eggs on leaf conceptual presentation in [Figure 1(A4)], where they are consumed by the nymphs just after the hatch(Kikuchietal,2011). Symbionts can also be established in hemipteran honeydew[Figure 1(A3)] and siphuncular fluids[Figure 1(A1)] that can persist on plants, where they might be transferred by other individuals(Darbyetal,2003).Rickettsia, one of the abundant endosymbionts carried by whiteflyB.tabaci, can inhabit phloem cells and spread all over the plant, and then infect a symbiont-free insect[Figure 1(D1)](Caspi-Flugeretal,2011).B.tabaciis extremely polyphagous species, but this plant facilitated horizontal transmission has been proved only in cotton currently. Symbionts present in gut biota of insects are closely related to microbes present in the environment, and the insect gut might be one of the continuums of habitats for them to exploit and in which they imitate(Grkovicetal,1995).
6 Plant pathogens as insect mutualists
Some pathogens and insect symbionts can use plants as an intermediary to do infection in new hosts, while pathogens of plant may use insects as a vector to transmit between plants. These may happen via crosstalk between signaling pathways engaged in plants reactions to insects and plant pathogens, specifically the salicylic acid and the jasmonic acid pathways(Kessleretal,2002;Pieterseetal,2007,2009;Dickeetal,2009). Various insects, comprising aphids, whiteflies, thrips and leafhoppers that can vector plant pathogenic fungi, bacteria or virus, perform healthier on, and are more attracted to, plants infested with the pathogens(Ebbertetal,2001;Kluthetal,2002;Fereresetal,2009).
The development of the novel symbioses can be leaded to different plant pathogens and consequently influence the significance of insect as a pest. TheDendroctonusvalensintroduced into China has acquired new native isolates of its associateLeptographiumprocerum. This new mutualism makes the beetle a considerably more severe pest in China than in its native distribution areas because the fungus modifies tree′s profile of the chemicals such that initiates many beetles towards host trees(Luetal,2010,2011).
Phylogenetic studies provide additional proof of close relationships between insect mutualists and plant pathogens.Spiroplasmahave species that are insect pathogens or mutualists, while some species have been recognized to be plant pathogens transmitted by psyllids(Gasparich,2010). The commonly circulated bacterial symbiontsArsenophonususually colonize other arthropods and insects but two species of this genus also found in plants as pathogens(Novákováetal,2009). For example,CandidatusA. phytopathogenicus can be transmitted vertically within the plant-hopperPentastiridiusleporinus, while it is a plant pathogen which can also be transferred between plants by the plant-hopper(Bressanetal,2009).
7 Insect symbionts influence plant physiology
In order to get self benefit herbivores have been testified to manage host plant physiology directly(Pieterseetal,2009). Herbivores can make different species-specific defenses in plants. They affect herbivore directly as well as indirectly by endorsing the natural enemies′ efficiency(Schoonhovenetal,2005;Pieterseetal,2009;Dickeetal,2010). There is overwhelming proof that insect-associated microbes benefit their hosts by interfering reaction in plant to attack of insects. For instance, in bark the mutualistic fungi and ambrosia beetles that attack wood help the insects in disabling tree resistance mechanisms(Paineetal,1997). Likewise, in theBactericercacockerelli, high concentrations of endosymbiontCandidatusLiberibacter psyllaurous associates with condensed expression of the plant pathways of defense directly in tomato(Casteeletal,2012).
In chewing insects, the most investigated chemical resisting plant defenses, an essential class N-acylamino acids, are present in insect saliva(Bonaventureetal,2011). The symbiotic flora of gut has potential to conquer the response of plant because 50% of 23 strains of symbiotic bacteria isolated from caterpillar′s gut have capacity to produce N-acylamino acids(Spitelleretal,2000). Another possible example consisted of leaf-mining moths prevent the residential area of the leaves on which they feed on senescing, leading to obvious ‘green islands’ produced by secreting cytokinins in autumnal colored leaves(Gironetal,2013). Recent studies have exposed that, in thePhyllonorycterblancardella, bacterial endosymbiont Wolbachia is involved in the cytokinins production(Kaiseretal,2010).
8 Insect symbionts suppress plant defense
Microorganisms associated with insects can also alter the insect-plant interactions. Insects and plants have some effects on their interface due to the presence of microorganisms. Some symbionts live inside the body of the insect and counter the defensive responses of plant by increasing enzymes production essential for insects.Dendroctonusponderosaeinhabits and kills conifers in America and expands its host range by climatic change(Raffaetal,2013). Pine trees produce terpenoids that are harmful to beetles. Recently metagenomics study reported thatD.ponderosaeis associated with microbial groups that are supplemented with genes that involve in terpene degradation as compared to other insects(Adamsetal,2013). Besides, the microbial community linked withD.ponderosaewere proved to digest monoterpenes and diterpene(Booneetal,2013). As another example,Diabroticavirgiferavirgiferahave symbiotic bacteria which facilitate to feed nonhost plants, e.g., soybean(Glycinemax)(Chuetal,2013). In such a situation, the activity of cysteine protease, a digestive enzyme, is increased which is counterattack against plant resistance due to existence of the gut microbes(Curzietal,2012).
Insect saliva has a direct association with the plant, and it can manipulate the biology of plant cells. Insect saliva contains various symbionts which can interfere with the consequences of insect-plant interaction.Leptinotarsadecemlineatahave many bacterial symbionts in its saliva that can be transmitted to plants during feeding[Figure 1(A1)]. The Flagellin result from thePseudomonassp. bacteria of this beetle conquers plant signaling pathway and make the plant susceptible for the beetle, which leads to weight increase of the beetle(Chungetal,2013). As another example, bacterial symbiont Wolbachia sp. of the Western corn rootworm make the maize plant less resistant to the insect(Barretal,2010). Besides, the GroEl derived from the primary aphid endosymbiontB.aphidicola, recognized in saliva of theMacrosiphumeuphorbiae, may reduce the aphid fecundity on tomato. However, while GroEL elicits plant defense reactions, aphids continue feeding on the plants, proposing that the secondary symbionts of aphids established strategies to suppress the GroEL defense reactions(Chaudharyetal,2014).
9 Conclusion and future perspectives
Insects have established very multifarious organism community accomplished of flexible variations to the usual challenges to which insect is exposed. Symbionts and their hosts can affect the suitability of each other. They together can be called holobionts with a hologenome encountering the obstacles during feeding related to nutrition, digestive and defensive processes. Insects have no or less alterable immune system, so the microbes are present in insects to interact with other factors. Insects with specialized feeding habitats have diverse symbiotic communities, such as termites and scarab beetles that feed on rich lignocellulose, complex polysaccharides and store food for longer period of time depend very much on symbionts as holobiont partners. Insect gut shows a huge diversity in morphology, food contents and physiochemical properties, which help to modify the microbiota within it, and the microbes contribute to insect adaptation in the environment. Many previous studies based on 16S rRNA sequencing of insect symbiotic microbes only focused on specific symbiont species, but we still know little about the exact diversity of symbiotic microorganisms and their functions, as well as genetic or genomic mechanisms underlying the functions in different insect lineages. Besides, in the future we should pay more attention to investigation of natural interactions between insects, plants and microbes in wild populations, because studies only restricted in laboratory usually can′t reveal the true diversity of interactions between symbionts and other partners as well as their functions. The more and more easy-to-use research technologies, such as high throughput 16S rRNA sequencing, metagenomics, and complete transcriptome, should help investigate these issues. Moreover, it′s important to understand how insects and plants differentiate between mutualistic symbionts and pathogens. From an applied science perspective, this may help manipulate the symbiotic microorganisms to provide beneficial functions to enhance food security and yield, as well as control destructive insect pests.
Acknowledgements:We would like to thank H. Zhang and Q. Liu for their help during the preparation of the manuscript.
