稻米脂肪与品质的关系及其调控
2020-07-23吴焱袁嘉琦张超张诚信陈天晔顾睿侯均昊周驰燕李国辉戴其根霍中洋许轲
吴焱 袁嘉琦 张超 张诚信 陈天晔 顾睿 侯均昊 周驰燕 李国辉 戴其根 霍中洋 许轲


摘要:脂肪是稻米第3丰富的重要营养组分,是稻米“能量棒”,是水稻生命活动所需能量的重要来源之一。但稻米中粗脂肪含量较少且提取困难,前人对于稻米脂肪的研究多集中于稻米脂肪与品质的关系以及稻米脂肪酸组成成分上,关于稻米脂肪含量和组分调控的研究较少。本文综述了水稻籽粒脂肪的种类及分布、合成和分解代谢及其调控途径,以及稻米脂肪含量及其脂肪酸组分与稻米品质尤其是稻米蒸煮食味品质的紧密联系和影响因素,展望了提高和改善稻米脂肪含量及其组分的技术途径,以期为稻米脂肪与品质协同改良提供帮助。
关键词:水稻;脂肪;品质;调控
中图分类号:S511.032文献标识码:A文章编号:1000-4440(2020)03-0769-08
The relationship between lipid and quality and its regulation in rice
WU Yan,YUAN Jia-qi,ZHANG Chao,ZHANG Cheng-xin,CHEN Tian-ye,GU Rui,HOU Jun-hao,ZHOU Chi-yan,LI Guo-hui,DAI Qi-gen,HUO Zhong-yang,XU Ke
(Jiangsu Key Laboratory of Crop Genetics and Physiology/Innovation Center of Rice Cultivation Technology in Yangtze Valley, Ministry of Agriculture/Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China)
Abstract: Lipid is the third most important nutrient component of rice, it is the "energy bar" of rice and the main provider of energy for rice life activities. The crude lipid content in rice is low, and the extraction is difficult. The previous studies on rice lipid mainly focused on the relationship between rice lipid and quality and composition of fatty acids in rice. There were few studies on the regulation of rice lipid content and composition. This paper reviewed the types and distribution of rice grain lipid, their synthesis and catabolism and their regulatory pathways, as well as the close relationship between rice lipid content and fatty acid components and rice quality, especially the cooking and eating quality of rice, and their influencing factors. The technical approaches for improving the lipid content and components of rice were prospected to provide help for the cooperative improvement of lipid and quality.
Key words:rice;lipid;quality;regulation
目前全世界大约有一半以上人的主食是稻米,水稻作为主要的世界性粮食作物,其作用不可忽视,而随着人们生活品质的不断提高,对于提高稻米品质的要求也日益迫切[1]。脂肪为人体生命活动提供能量[2]。稻米的脂肪含量是比较低的,其中多数是不饱和脂肪酸或淀粉-脂复合物,这些物质影响着稻米的品质,且对于心血管疾病具有良好的预防效果,营养价值较高[3-4]。一般来说,脂肪含量较高的稻米食味品质较好[5]。脂肪对于提升稻米食味品质具有積极影响,较高脂肪含量的稻米蒸煮后,各项食味评价指标表现均较佳[6]。但在稻米的储藏过程中,脂肪在环境作用及相关酶的影响下,会发生酸变,最终导致稻米陈化,品质下降[7]。
脂肪,即甘油三酯(TAG),由甘油和三分子脂肪酸组成。脂肪酸是甘油三酯的主要成分,其合成包含很多复杂的生化反应,目前植物脂肪酸代谢途径已经越来越明晰,通过脂肪合成过程中相关酶及其基因来调控脂肪含量,最终实现调控稻米品质已经成为可能[8]。本文通过分析稻米脂肪的分布、分类、合成代谢、调控机制等,明确脂肪与稻米品质之间的关系,为稻米脂肪以及调控的深入研究提供帮助。
1稻米脂肪的分类及分布
脂肪是稻米的第3大物质组分,在糙米中粗脂肪含量在4%以内,精米中不足1.5%。稻米脂肪大多分布在胚中,其次是种皮和糊粉层,胚乳中含量最少[9];胚乳中脂肪分布不均匀,越往内部,淀粉晶体分布越紧密,脂肪含量越低[10]。
可以将稻米中的脂类分为2类:溶于热的水饱和正丁醇溶液的脂类为淀粉脂类(SL),非极性脂类、糖脂和磷脂这些都属于淀粉脂类,而最主要的组成部分是磷脂[11],其中溶血卵磷占很大比例(约85%)。需要冷的水饱和正丁醇溶液溶解的是非淀粉脂类(NSL),圆球体和与蛋白体结合的脂肪属于此类。糯稻淀粉脂类(SL)含量明显低于非糯稻,非淀粉脂类(NSL)含量则明显高于非糯稻。在非糯稻中,糙米直链淀粉含量升高,淀粉脂类(SL)含量也随之增加[12]。从结构上可以将谷糠中的脂类分为3种:以甘油和脂肪酸为共同结构的甘油脂类,以有固醇为共同点的固醇脂类和以有鞘氨醇和脂肪酸为共同点的鞘脂类。
2稻米脂肪的合成与分解代谢
稻米脂肪酸的主要组分是油酸、亚油酸、棕榈酸,约占总脂肪酸含量的90%。其他脂肪酸含量如亚麻酸、硬脂酸和花生酸含量均不高[13],主要为不饱和脂肪酸。
2.1脂肪的合成代谢及调控
2.1.1脂肪的合成代谢图1为植物种子脂肪及脂肪酸生物合成途径[14]。植物脂肪合成部位为叶绿体和前质体,合成原料由葡萄糖代谢提供。脂肪酸代谢在质体和内质网中发生。首先,丙酮酸通过糖酵解途径生成后,在丙酮酸脱氢酶复合物的作用下,被氧化为乙酰辅酶A。16C及以下饱和脂肪酸的合成采用从头合成途径。以软脂酸合成为例,线粒体内的乙酰辅酶A通过柠檬酸-丙酮酸循环出线粒体,在乙酰辅酶A羧化酶的作用下合成丙二酸单酰辅酶A。之后1个乙酰辅酶A和7个丙二酸单酰辅酶A在脂肪酸合成酶的催化下依次经历启动、装载、缩合、还原、脱水、再还原等6种反应,最终合成软脂酸[15]。16C以上的饱和脂肪酸,如硬脂酸、棕榈酸,需要在光滑内质网和线粒体中在脂肪酸-碳链延伸酶系统的催化作用下形成更长的碳链饱和脂肪酸[16]。
不饱和脂肪酸的生物合成是饱和脂肪酸去饱和合成,去饱和作用在微粒体中进行。脂酰辅酶A和3-磷酸甘油在甘油磷酸转酰基酶催化作用下缩合生成磷脂酸,同时磷酸二羟丙酮也与脂酰辅酶A反应生成脂酰磷酸二羟丙酮,然后还原生成溶血磷脂酸,溶血磷脂酸和脂酰辅酶A作用可生成磷脂酸。磷脂合成的中间产物为磷脂酸。磷脂酸磷酸酶催化磷脂酸生成二酰甘油和磷酸,二酰甘油与另一分子的脂酰辅酶A缩合生成三酰甘油[17]。
2.1.2脂肪合成的调控
2.1.2.1关键酶及其基因的调控乙酰辅酶A羧化酶(ACCase)在脂肪酸合成途径中起着关键作用,其催化的反应可以限制脂肪酸合成速度,并控制碳流进入脂肪酸合成途径[18]。因此,调节ACCase活性及其基因表达可以作为调控脂肪酸合成的一个重要切入点。ACCase可以分为同质型和异质型两大类,其中同质型ACCase存在2种不同基因形式Acc-1和Acc-2[19]。研究结果表明水稻脂肪酸合成途径中的ACCase属于同质型多功能酶,且其编码基因为Acc-2[20]。目前ACC基因的全長序列已经在小麦、玉米等植物中被解析。提高ACCase活性能够促进脂肪酸的合成,相反,ACCase活性降低,脂肪酸含量也相应减少,这一点在表达生物素羧基转运的转基因植物中得到了证实[21]。把从拟南芥中克隆的Acc-1基因转入到油菜中,发现油酸含量提高[22]。但关于水稻ACC基因克隆研究的详细报道还比较少见[23]。在底物竞争假设中,在磷酸烯醇式丙酮酸羧化酶(PEPCase)的参与下,丙酮酸合成草酰乙酸,进入蛋白质代谢,如果抑制PEPC基因的表达,可以促进更多的丙酮酸用于脂肪酸的合成,进而增加稻米脂肪含量[24]。但也有研究者发现,将从玉米中提取的PEPC基因直接导入水稻中,光合效率显著提高,较对照组脂肪含量有较大幅度提高[25]。
脂肪酸合酶复合体在脂肪酸合成过程中是不可缺少的,它由酰基载体蛋白、β-酮脂酰-ACP脱水酶、β-酮脂酰-ACP合酶、β-酮脂酰-ACP-还原酶、烯脂酰-ACP还原酶、脂酰-ACP硫脂酶等部分构成,这些酶参与了脂肪酸合成途径中的不同反应。目前,酰基-ACP硫酯酶(FAT)已经从拟南芥[26]、向日葵[27]中得到纯化并且克隆了其编码基因。研究结果表明,将从拟南芥中克隆到的FATB1基因转入到其他植物种子中,棕榈酸含量增加[28];若抑制FATB1表达,会减少棕榈酸含量[29]。
将脂酰链转移到甘油分子的碳骨架上是脂肪合成的关键步骤,二酰甘油酰基转移酶(DGAT)[30] 在这步骤中起不可忽视的作用,且该酶的QTL已经在水稻中被检测出[31]。目前已发现DGAT1、DGAT2、WS/DGAT和可溶性DGAT这4类DGAT。研究发现在大豆[32]和玉米[33]中通过异位表达DGAT基因能够显著提高种子脂肪含量,同时发现DGAT1和DGAT2受植物品种以及不同生育阶段的影响,并起着不同的作用,进而合成不同组分的TAG[34]。
2.1.2.2基因工程的调控饱和脂肪酸对人体心脑血管有不利影响,因此降低稻米饱和脂肪酸含量,提高不饱和脂肪酸含量具有重要意义[35]。稻米脂肪中饱和脂肪酸主要是棕榈酸,降低棕榈酸除了采用常规栽培方法外还可通过分子遗传调控技术,促进参与聚合反应的酶基因的表达,降低棕榈酰硫酯酶活性,从而降低棕榈酰基-ACP含量。主要有3种方法:一是提高β-酮脂酰-ACP合酶基因的表达水平,对于降低棕榈酸含量有直接效应;二是降低棕榈酰硫酯酶活性,在植物中转入活性更强的酰基硫酯酶是很有效的做法[36];三是提高脱饱和酶的活性,这一点可以通过促进脱饱和酶基因的表达来实现。脱饱和基因的表达有利于增加不饱和脂肪酸的含量且降低饱和脂肪酸含量。
导致稻米在贮藏过程中陈化的主要原因是:含有多个双键的不饱和脂肪酸容易被氧化。因此降低多不饱和脂肪酸含量是延缓稻米陈化的重要途径。前人多采用去掉双键的方法,但该方法最大缺点是会使顺式双键变成反式,而食用大量反式不饱和脂肪酸会增加冠心病的发病几率。因此,通过调控硬脂酸、油酸等脱饱和酶活性以降低多不饱和脂肪酸含量是更加可行的途径。将植物种子内硬脂酸和油酸的脱饱和酶基因反义表达,降低酶活性,可以提高硬脂酸和油酸的含量,尤其是油酸含量得到大幅度提高[37]。研究结果表明抑制脂肪酸脱氢酶(FAD2)基因的表达,不仅能减少对人体健康不利的棕榈酸以及引起稻米陈化的亚油酸含量,还能提高稻米油酸含量[38]。
2.1.2.3植物激素与环境因子的调控植物激素脱落酸(ABA)能作用于种子的多种生理过程,对于脂肪含量调节有直接影响,可对脂肪的积累有促进作用并且抑制脂肪的分解[39]。
影响植物脂肪含量的环境条件有很多,水分、温度、空气、光照以及矿质元素等都会对其产生一定程度的影响。产生ATP和NADH(NADPH)的必要条件之一是光,因此,光对植物脂肪合成有重要作用。研究发现,在有光条件下油菜[40]、大豆[41]等植物种子含油量增加。水稻灌浆时期若处于弱光条件下脂肪含量降低,但不饱和脂肪酸含量增加[42]。遮光条件对大豆脂肪酸含量和组成的影响因大豆品质的不同而不同[43]。
温度对于脂肪的合成及积累也有很大影响。在灌浆结实期,磷脂酸磷酸酯酶(PPase)、6-磷酸葡萄糖脱氢酶(G6PD)和PEPCase活性受温度影响较大,进而影响脂肪的合成,且高温会抑制PPase和G6PD活性但增加PEPCase活性,进而导致稻米脂肪含量降低[42]。同时也发现日间高温不利于稻米脂类的积累[44],但夜间高温有利于稻米脂类的合成[45],成熟期稻米不饱和脂肪酸含量则随着温度升高而降低[46]。通过控制水稻不同生育时期水分条件发现,抽穗前控水会增加籽粒的脂肪酸含量,抽穗后1~10 d控水会降低脂肪酸含量,表明水分与稻米脂肪酸含量密切相关且受生育时期的影响[47]。在肥料施用方面,随着氮、磷、钾肥料施用量的增加,玉米籽粒饱和脂肪酸含量、不饱和脂肪酸含量呈先增后减的趋势,氮、磷、钾水平为复合肥450 kg/hm2、尿素600 kg/hm2时饱和脂肪酸含量和不饱和脂肪酸含量均达到最高[48]。而稻米粗脂肪含量随施氮量的增加也呈现出先增后减的趋势[49],且钾肥和磷肥的施用能够显著增加稻米脂肪含量,可见肥料的使用对于稻米脂肪的合成有一定的影响[50]。
2.2稻米脂肪的分解代谢
生物利用三酰甘油作为能源的第一步是利用脂肪酶水解三酰甘油,产生甘油和脂肪酸。甘油和脂肪酸被氧化,释放出的化学能量被生物体用于各种生理活动。甘油在甘油激酶的作用下生成3-磷酸甘油,再经磷酸甘油脱氢酶,在NAD+参与下,生成磷酸二羟丙酮及NADH+H。磷酸二羟丙酮可以经糖酵解过程转变为丙酮酸,再进入三羧酸循环氧化。
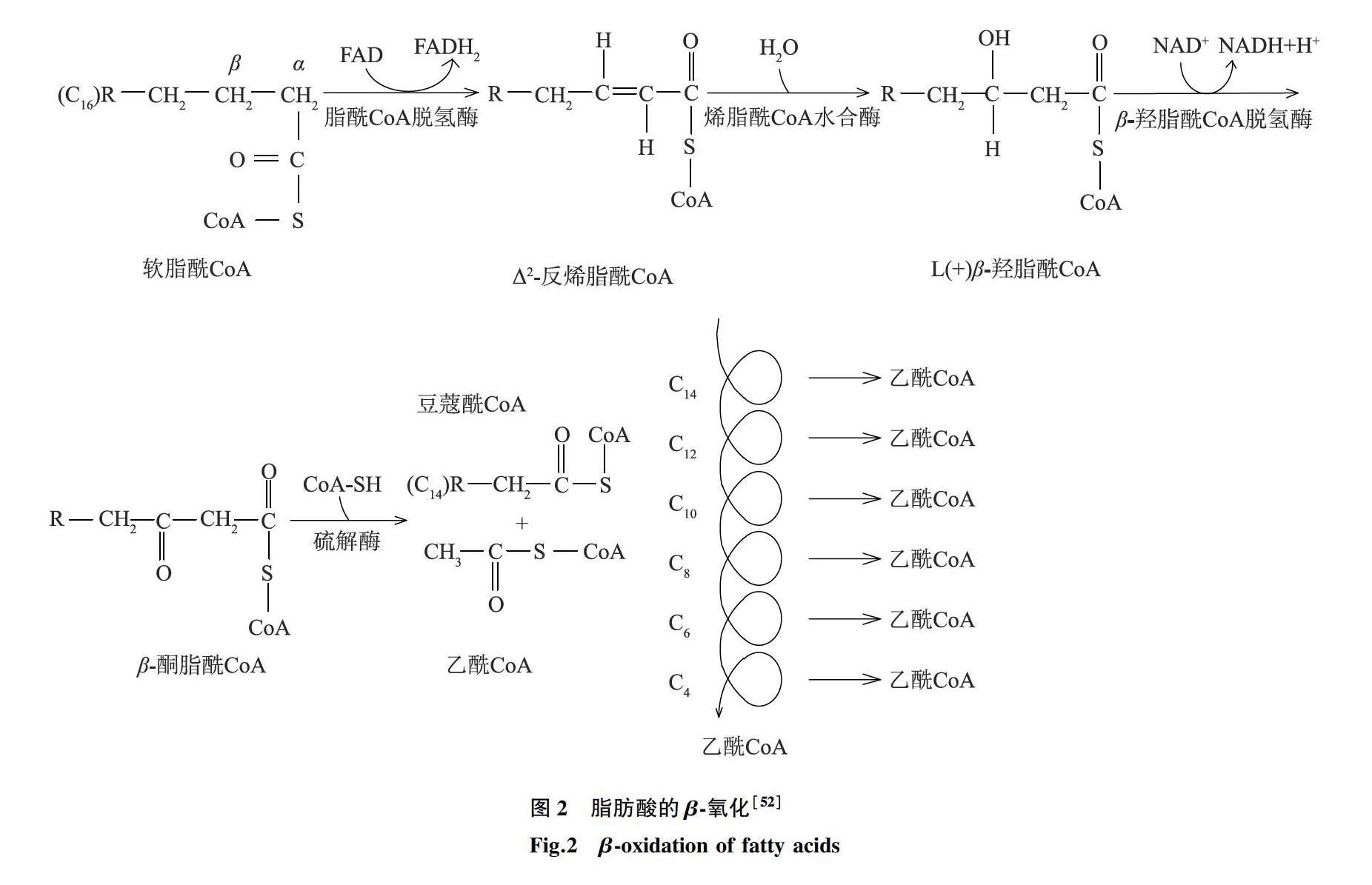
脂肪酸氧化可分为β-氧化、α氧化、ω氧化等。脂肪酸降解时,从羟基端β-C原子开始,经脱氢、水化、再脱氢、硫解4步反应生成一个二碳化合物(乙酰辅酶A)和比原脂肪酸少2个C原子的脂酰辅酶A,这一过程称β-氧化[51]。脂肪酸经过活化后生成脂酰辅酶A,以软脂酰辅酶A为例的β-氧化具体过程见图2[52]。不饱和脂肪酸的氧化、脂肪酸活化与转运同饱和脂肪酸。但单不饱和脂肪酸需要异构酶进行β-氧化,而多不饱和脂肪酸则需要异构酶和还原酶的参与才能进行β-氧化。稻米中含奇数碳原子的脂肪酸进行β-氧化后,产物是丙酰辅酶A和乙酰辅酶A。这与含偶数碳原子的脂肪酸氧化后产物不同,丙酰辅酶A 的代谢见图3[52]。
脂肪酸除进行β-氧化外,还进行α-氧化、ω-氧化。在α-氧化途径中,长链脂肪酸的α碳在单加氧酶的催化下氧化为羟基,形成α-羟脂酸,经过进一步脱羧氧化转变为比原脂肪酸分子少一个碳原子的脂肪酸。同时,脂肪酸的末端甲基可氧化为羟脂酸,然后氧化为α,ω-二羧酸进行β氧化,这就是ω-氧化[52]。
3脂肪含量与稻米品质的关系
3.1脂肪含量与稻米外观品质的关系
外观品质是稻米商业价值的重要参考之一,是指糙米或者精米的外表物理特性,包含垩白、透明度、粒形等评价指标[53]。目前发现脂肪含量高的稻米表现出油质鲜明的特征[54],且脂肪含量与稻米粒长、粒宽、长宽比呈显著的正相关关系[55]。
3.2脂肪含量与蒸煮食味品质的关系
稻米蒸煮食味品质是指在蒸煮与食用过程中稻米表现出来的理化与感官特性。大量研究结果表明在一定范围内,脂肪含量越高,米饭光泽越好,米饭适口性与香气也越佳[5]。
3.2.1脂肪含量与稻米蒸煮品质的关系衡量蒸煮品质的主要指标包括胶稠度、直链淀粉含量、淀粉黏滞特性、糊化特性、米粒的延伸性等。直链淀粉含量和胶稠度反映米饭的物理特性。淀粉的糊化特性通过差示扫描量热分析仪(DSC)测定值反映,黏滞特性通过快速黏度分析仪(RVA)测定值表现。
虽然稻米中脂类含量很少,但在蒸煮品质上起着很重要的作用。前人研究发现脂肪对淀粉糊化有重要影响,虽然在淀粉糊化过程中,直链淀粉只起到稀释的作用,但是在天然淀粉中,直链淀粉与脂类中磷脂和糖脂形成的复合物起到抑制淀粉膨润[56],提高糊化温度的作用[57],进而影响稻米的蒸煮品质[58]。目前发现稻米粗脂肪含量与直链淀粉含量相关性不一致[59-60],但可确定的是其中淀粉脂类(SL)含量与直链淀粉含量呈显著正相关关系。而稻米脂肪含量与胶稠度则表现出极显著正相关关系[61]。比较脱脂后米粉RVA谱特征值发现峰值黏度、热浆黏度、冷胶黏度比脱脂前均变大,可以看出黏度与脂肪含量呈极显著负相关关系,这是因为脂肪具有润滑特性[62]。崩解值与脂肪含量则呈显著负相关关系。而崩解值、消减值、回复值与脂肪中的淀粉脂类呈极显著负相关关系[63]。但陈米的糙米米粉由于脱去了非淀粉脂类,消除或减弱了脂类与淀粉相互作用,其最高黏度值、最终黏度值有所下降,对淀粉糊化造成了影响[64]。陈米精米米粉脱脂后其最高黏度值也呈下降趋势[65],这可能是由于储藏一段时间后直链淀粉已经老化,支链淀粉开始发生老化,而淀粉膨润是支链淀粉的特性,因此,淀粉的膨润力下降导致最高黏度值下降。研究脱脂精米米粉DSC热谱曲线发现较脱脂前热焓值显著降低[66]。糙米米粉脱去非淀粉脂类后DSC热谱曲线中峰值温度明显下降,可以看出非淀粉脂类对于淀粉糊化影响较大[67]。在储藏过程中,稻米陈化与脂肪酸组分变化有很大关系,在储藏一段时间后发现非淀粉脂类和淀粉脂类的总含量保持不变,但非淀粉脂类含量及其组分变化活跃,主要是非淀粉脂类中的亚油酸和亚麻酸氧化产生丙醛、丙酮、戊烷和已烷,使得陈米中的羰基化合物含量增加。这主要是因为非淀粉脂类大多存在于脂解酶类较多的地方,而淀粉脂类多与直链淀粉形成复合物,处于稳定的状态[68]。
3.2.2脂肪含量与稻米食味品质的关系稻米的食味品质主要是米饭给人留下的感官印象,包括颜色、香气、黏性、弹性、综合口感以及光泽度、米粒的完整性等。通常有光泽、气味清香、质地柔软有弹性、适口性佳的米饭被认为是品质优良的米饭。
目前研究发现,稻米脂肪对稻米的食味品质有重要作用[69],在一定范围内,脂肪含量越高,米粒完整性更佳,米饭更油亮,光泽度越高,米饭的适口性和香味也更好[70-71]。且研究发现稻米脂肪中不饱和脂肪酸对米饭适口性具有积极影响[72]。脂肪对于米饭的质构特性即米饭的黏度、弹性、硬度、平衡度也有一定的影响[73]。前人研究发现稻米的粗脂肪含量与米饭的硬度呈负相关关系,与黏度呈正相关关系[74]。因此,稻米脂肪含量对于提高稻米的食味品质有积极影响。此外,脂类能够保护直链淀粉不会在热加工过程中因高温挤压而发生裂解,有利于提高膨化食品口感[75]。研究发现,随着脂肪酸添加量的增加,米饭硬度变大,黏度降低,油酸较其他脂肪酸对降低米饭黏性有更显著作用。这是由于随着淀粉-脂复合物的积累,淀粉內部结构被充实,稻米变得紧实[76]。
3.3脂肪含量與稻米淀粉性质的关系
脂肪对稻米淀粉性质产生影响主要是通过形成淀粉-脂复合物。淀粉-脂复合物的形成使淀粉原有的糊化性质、老化性质、消化性质等均发生改变[77]。脂肪能够抑制淀粉颗粒的溶胀,提高糊化温度[78],从而限制淀粉的糊化进程[79]。此外,淀粉-脂复合物结晶与淀粉老化存在竞争关系,且结晶比老化更容易发生,因此淀粉-脂复合物对于延缓淀粉老化有积极作用[80]。另外,脂质本身就有利于维持淀粉凝胶的网状结构,这一特性也抑制了淀粉老化[81]。同时,淀粉-脂复合物能够提高淀粉的抗消化性质主要有两方面原因:一是能够阻止消化酶进入淀粉内部[82],减少酶与淀粉络合的机会[83];二是淀粉-脂复合物本身的抗消化性就比淀粉高[84]。
4展望
目前,水稻的发展追求已经偏向于优质高产,提高水稻品质也成为稻作研究的一大热点。脂类作为稻米中含量第3丰富的营养物质,含量虽不高,但对其品质的影响很大。因此,合理调控稻米脂类含量及组分对优质稻米育种和功能性优质稻米开发具有重要意义。
相关研究结果表明,稻米中棕榈酸为饱和脂肪酸,大量进食易得心脑血管疾病;亚油酸和亚麻酸会导致稻米陈化,影响稻米品质,且其为多不饱和脂肪酸,对人体心脏也有不利影响。因此,当前调控脂肪酸的主要任务是在提高脂肪酸含量的基础上,降低饱和脂肪酸含量,增加单不饱和脂肪酸含量。通过基因工程或者环境因子、植物激素等调控途径来调控脂肪酸组分及含量是可行的基础途径。脂肪的合成和分解代谢需要一系列酶基因的表达,且环境因素以及种子自身发育情况都对脂肪合成和分解有较大影响[85]。目前对脂肪合成基因的转录调控因子以及参与酶的协同调控还需要更全面深入研究。
由于稻米脂肪含量少、提取难等因素,以水稻为试验材料探寻水稻脂肪酸调控的研究还很少。前人关于脂肪调控的研究多集中于高油植物,虽然得出了很多成功的调控途径,但在水稻上尚需要进一步验证。随着对种子油及脂肪酸研究的不断深入,稻米脂肪调控终将越来越完善,获得更好吃的优质高产稻米这一目标终将会实现。
参考文献:
[1]于永红,朱智伟,程方民. 稻米的脂肪[J]. 中国稻米,2006(3):12-13.
[2]陈金波. 植物种子中脂肪酸代谢调控基因工程研究[J]. 现代农业科学,2009, 16(5):20-21.
[3]李懿璞. 稻米脂肪含量的研究进展[J]. 农技服务,2010,27(3):399-400.
[4]MORRISON W R, MILLIGAN T P, AZUDIN M N. A relationship between the amylose and lipid contents of starches from diploid cereals[J]. Journal of Cereal Science,1984,2(4):257-271.
[5]伍时照,黄超武,欧烈才,等. 水稻品种种性研究Ⅲ.水稻品种品质性状的研究[J]. 中国农业科学,1985,18(5):1-7.
[6]蔡洪法,刘新录. 中国稻米品质区划及优质栽培[M]. 北京:中国农业出版社,2002.
[7]张瑛,吴先山,吴敬德,等. 稻谷储藏过程中理化特性变化的研究[J]. 中国粮油学报,2003,18(6):20-28.
[8]BEWLEY J D, BLACK M. Seeds: physiology of development and germination[J]. Experimental Agriculture,1995,31(4):445-515.
[9]WANG H L, WAN X Y, BI J C, et al. Quantitative analysis of fat content in rice by near-infrarde spectroscopy technique[J]. Cereal Chemistry,2006,83(4):402-406.
[10]ZHOU Z, ROBARDS K, HELLIWELL S, et al. Composition and functional properties of rice[J]. International Journal of Food Science & Technology, 2002, 37(8):849-868.
[11]JULIANO B O. Criteria and tests for rice grain qualities[M]/ /Juliano B O. Rice: Chemistry and technology. Minnesota: AmericanAssociation of Cereal Chemists,1985.
[12]刘保国,成萍,卢季昌,等. 水稻籽粒脂肪及脂肪酸组分的分析[J]. 西南农业大学学报,1992, 14(3):275-277.
[13]王瑞,施莉莉,张欣,等. 几个稻米品种(系)中脂肪酸含量的研究[J]. 安徽农业科学,2011,39(14):8592-8593.
[14]王计平,史华平,李润植. 植物种子油合成的调控与遗传修饰[J]. 植物遗传资源学报,2006,7(4):488-493.
[15]徐瑞新,王洁,任国勇,等. 植物脂肪酸合成及其在基础抗性和生物固氮中的作用研究进展[J].湖北农业科学,2016,55(22):5729-5734.
[16]石东乔,周奕华,陈正华. 植物脂肪酸调控基因工程研究[J]. 生命科学,2002,14(5):291-295,317.
[17]卢善发. 植物脂肪酸的生物合成与基因工程[J]. 植物学通报,2000,17(6):481-491.
[18]SLABAS A R, FAWCETT T. The biochemistry and molecular biology of plant lipid biosynthesis[J]. Plant Molecular Biology,1992,19(1):169-191.
[19]CRONAN J E, WALDROP G L. Multi-subunit acetyl-CoA carboxylases[J]. Progress in Lipid Research,2002, 41(5):407-435.
[20]GENGENBACH B G, SOMERS D A, WYSE D L, et al. Transgenic plants expressing maize acetyl coa carboxylase gene and method of altering oil content: US,6222099[P].2001-04-24.
[21]任波,李毅. 大豆種子脂肪酸合成代谢的研究进展[J]. 分子植物育种,2005,3(3):301-306.
[22]ROESLER K. Targeting of the arabidopsis homomeric acetyl-coenzyme a carboxylase to plastids of rapeseeds[J]. Plant Physiology,1997,113(1):75-81.
[23]李洁琼,郑世学,喻子牛,等. 乙酰辅酶A羧化酶:脂肪酸代谢的关键酶及其基因克隆研究进展[J]. 应用与环境生物学报,2011,17(5):753-758.
[24]吴关庭,郎春秀,胡张华,等. 应用反义PEP基因表达技术提高稻米脂肪含量[J]. 植物生理与分子生物学学报,2006,32(3):339-344.
[25]张边江,唐宁,陈全战,等. 转C4基因水稻籽粒产量及品质分析[J]. 西北农业学报,2017,26(2):210-215.
[26]DORMANN P, VOELKER T A, OHLROGGE J B. Cloning and expression in escherichia coli of a novel thioesterase from arabidopsis thaliana specific for long-chain acyl-acyl carrier proteins[J]. Archives of Biochemistry and Biophysics,1995,316(1):612-618.
[27]SERRANO-VEGA M J, GARCS R, MARTNEZ-FORCE E. Cloning, characterization and structural model of a FatA-type thioesterase from sunflower seeds (Helianthus annuus L.)[J]. Planta, 2005, 221(6):868-880.
[28]DORMANN P, VOELKER T A, OHLROGGE J B. Accumulation of palmitate in arabidopsis mediated by the acyl-acyl carrier protein thioesterase FATBl[J]. Plant Physiology,2000,123(2):637-644.
[29]BONAVENTURE G, SALAS J J, POLLARD M R, et a1.Disruption of the FATB gene in arabidopsis demonstrates an essential role of saturated fatty acids in plant growth[J]. Plant Cell,2003,15:1020-1033.
[30]JAKO C, KUMAR A, WEI Y, et al. Seed-specific over-expression of an arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight[J]. Plant Physiology,2001, 126(2):861-874.
[31]王海莲. 稻米脂肪含量QTL分析及二酰甘油酰基转移酶基因的克隆[D].南京:南京农业大学,2007.
[32]LARDIZABAL K, EFFERTZ R, LEVERING C, et al. Expression of umbelopsis ramanniana DGAT2A in seed increases oil in soybean[J]. Plant Physiology,2008,148(1):89-96.
[33]ZHENG P, ALLEN W B, ROESLER K, et al. A phenylalanine in DGAT is a key determinant of oil content and composition in maize[J]. Nature Ge-netics,2008,40(3):367-373.
[34]OAKES J, BRACKENRIDGE D, COLLETTI R, et al. Expression of fungal diacylglycerol acyltransferase2 genes to increase kernel oil in maize[J]. Plant Physiology,2011,155(3):1146-1157.
[35]周奕华,陈正华.植物种子中脂肪酸代谢途径的遗传调控与基因工程[J]. 植物学通报,1998,15(5):17-24.
[36]YADAV N, WIERZBICKI A, KNOWLTON S, et al. Genetic manipulation to alter fatty acid profiles of oilseed crops[J]. Current Topics in Plant Physiology, 1993,9:60-66.
[37]KNUTZON D S, THOMPSON G A, RADKE S E, et al. Modification of brassica seed oil by antisense expression of a stearoyl-acyl carrier protein desaturase gene[J]. Proceedings of the National Academy of Sciences USA,1991,89(7):2624-2628.
[38]ZAPLIN E S, LIU Q, LI Z , et al. Production of high oleic rice grains by suppressing the expression of the OsFAD2-1 gene[J]. Functional Plant Biology,2013,40(10):996.
[39]YANG Y, YU X C, SONG L F, et al. ABI4 activates DGAT1 expression in arabidopsis seedlings during nitrogen deficiency[J]. Plant Physiology,2011,156(2):873-883.
[40]GOFFMAN F D, ALONSO A P, SCHWENDER J, et al. Light enables a very high efficiency of carbon storage in developing embryos of rapeseed[J]. Plant Physiology,2005,138(4):2269-2279.
[41]RUUSKA S A, SCHWENDER J, OHLROGGE J. The capacity of green oilseeds to utilize photosynthesis to drive biosynthetic processes[J]. Plant Physiology,2004,136(1):2700-2709.
[42]许光利. 稻米脂类对品质的影响及脂类代谢对高温弱光的响应[D]. 雅安:四川农业大学,2017.
[43]刘兵,王程,金剑,等. 生殖生长期遮阴对大豆脂肪酸积累动态的影响[J]. 大豆科学,2009,28(1):53-58.
[44]曾凯,居为民,周玉,等. 高温逼熟等级对早稻品质与产量特征的影响[J]. 中国农学通报,2011,27(30):120-125.
[45]COOPER N T W, SIEBENMORGEN T J, COUNCE P A. Effects of nighttime temperature during kernel development on rice physicochemical properties[J]. Cereal Chemistry, 2008, 85(3):276-282.
[46]KITTA K, EBIHARA M, IIZUKA T, et al. Variations in lipid content and fatty acid composition of major non-glutinous rice cultivars in Japan[J]. Journal of Food Composition and Analysis,2005,18(4):269-278.
[47]郑桂萍,刘沐江,刘丽华,等. 稻米脂肪酸与食味的关系及土壤水分的调控效应[J]. 中国生态农业学报,2009,17(6):1149-1155.
[48]黄清梅,江鲁华,赵自仙,等. 不同施肥处理对玉米的氨基酸组分及脂肪酸组分的影响[J]. 西南农业学报,2013,26(5):2062-2065.
[49]顾丹丹. 粳米脂肪含量的氮素效應及其与米粉理化特性的关系研究[D]. 南京:南京农业大学,2011.
[50]朱朋波,徐大勇,方兆伟,等. 氮磷钾肥施用量对稻米粗脂肪含量的影响[J]. 江苏农业科学,2006(6):395-396.
[51]徐靖,李辉亮,朱家红,等. 植物脂肪酸β-氧化的研究进展[J]. 生物技术通讯,2008,19(1):141-144.
[52]张丽萍,杨建雄. 生物化学简明教程[M].北京:高等教育出版社,2015.
[53]王琦. 粳稻蒸煮食味品质形成的理化基础研究[D]. 南京:南京农业大学,2016.
[54]祁祖白,李宝健,杨文广,等. 水稻籽粒外观品质及脂肪含量的遗传研究[J]. 遗传学报,1983,10(6): 452-458.
[55]陈建国,朱军,减荣春. 籼粳亚种间杂交稻米脂肪的遗传分析[J]. 热带亚热带植物学报,1998,6(4):347-351.
[56]陈银基,鞠兴荣,董文,等. 稻谷中脂类及其储藏特性研究进展[J]. 食品科学,2012,33(13):320-323.
[57]蔡丽明,高群玉.淀粉-脂类复合物的研究现状及展望[J].粮油加工,2007(2):85-87.
[58]LARSSON K. Inhibition of starch gelatinization by amylase lipid complex formation[J]. Starch-Strke,1980,32(4): 125-126.
[59]江谷驰弘,雷小波,兰艳. 粳稻脂肪含量对稻米品质的影响[J]. 华南农业大学学报,2016,37(6):98-104.
[60]JULIANO B O. Rice chemistry and technology[M]. St Paul Minnesota USA:AACC Inc,1985.
[61]于永红,周鹏,段彬伍,等. 水稻脂肪含量分布及与食味品质的相关性分析[J].浙江农业科学,2007(6):669-671.
[62]吴洪恺,刘世家,张文伟,等. 稻米脂肪与米粉RVA谱特征的关系分析[J]. 江苏农业学报,2009,25(3):464-468.
[63]顾丹丹,刘正辉,刘杨,等. 粳稻精米脂肪含量和组分对蒸煮品质的影响及其对氮素的响应[J].作物学报, 2011,37(11): 2001-2010.
[64]PEREZ C M, JULIANO B O. Texture changes and storage of rice[J]. Texture Studies,1981(12):321-333.
[65]张守文. 大米陈化过程中的质量变化及品质改良的研究[J]. 中国粮油学报,1997,12(1):10-15.
[66]刘奕,徐海明,程方民,等. 稻米脱脂与未脱脂米粉的DSC热力曲线和RVA特征值比较[J]. 浙江大学学报(农业与生命科学版),2005,31(5):518-523.
[67]CHAMPAGNE E T. Effect of degree milling and lipid removal on starch gelatinization in the brown rice kernel[J]. Cereal Chem,1990,67(6):570-574.
[68]YASUMASTU K, MORITAKA S. Fatty acid compositions of rice lipid and their changes during storage[J]. Agric Biolchem,1964(28):257-267.
[69]ZHOU Z, ROBARDS K, HELLIWELL S, et al. Composition and functional properties of rice[J]. International Journal of Food Science & Technology, 2002, 37(8):849-868.
[70]LIU Y B. Effects of fat content of rice on flavor quality[G]. Fukui, Japan: Proceeding of the 2nd Asian Crop Science Conference, 1995.
[71]賀萍,陈竞适,张喻,等. 大米蛋白质和脂肪含量对鲜湿米粉品质的影响[J]. 湖南农业科学,2017(1):73-76,80.
[72]YOON M R, KOH H J, KANG M Y. Variation of properties of lipid components in rice endosperm affected on palatability[J]. Journal of the Korean Society for Applied Biological Chemistry, 2008, 51(3):207-211.
[73]毕雪,张敏,周琦,等. 添加食用油对米饭食味影响的研究[J]. 食品科学技术学报,2018,36(6):39-50.
[74]CAMERON D K, WANG Y J. A better understanding of factors that affect the hardness and stickiness of long-grain rice[J]. Cereal Chemistry, 2005, 82(2) :113-119.
[75]刘奕,程方民. 稻米中蛋白质和脂类与稻米品质的关系综述[J]. 中国粮油学报,2006,21(4):6-10,24.
[76]毕雪,张敏,周琦,等. 脂肪酸对米饭食味的影响[J]. 食品科学, 2019,40(24):8-14.
[77]郭宏伟,许秀颖,赵城彬,等. 淀粉-脂质复合物的研究进展[J]. 食品工业,2018,39(6):237-240.
[78]KAUR K, SINGH N. Amylose-lipid complex formation during cooking of rice flour[J]. Food Chemistry, 2000, 71(4):511-517.
[79]CHEN X, HE X, ZHANG B, et al. Structure, physicochemical and, in vitro, digestion properties of ternary blends containing swollen maize starch, maize oil and zein protein[J]. Food Hydrocolloids, 2017,25(4) :88-95.
[80]刘延奇,秦令祥,吴史博,等.淀粉-脂质复合物的相互作用及研究进展[J].食品科技,2009,34(7):209-213.
[81]李雨露. 莲子淀粉老化及脂质抗老化机理的研究[D].南昌:南昌大学,2015.
[82]AI Y, HASJIM J, JANE J L. Effects of lipids on enzymatic hydrolysis and physical properties of starch[J]. Carbohydrate Polymers, 2013, 92(1):120-127.
[83]OKUMUS B N, TACER-CABA Z, KAHRAMAN K, et al. Resistant starch type V formation in brown lentil ( Lens Culinaris Medikus) starch with different lipids/fatty acids[J]. Food Chemistry, 2018, 240:550-558.
[84]CUI R, OATES C G. The effect of amylose-lipid complex formation on enzyme susceptibility of sago starch[J]. Food Chemistry, 1999, 65(4):417-425.
[85]周国雁,隆文杰,陈丹,等.袋装临沧铁壳麦种子生活力及α-淀粉酶、脂肪氧化酶活性与表达量分析[J].南方农业学报,2018,49(11):2273-2281.
(责任编辑:张震林)
