低温胁迫对不同杂交水稻组合苗期耐冷性及生理生化指标的影响
2020-07-20沈航陈娇黎佰杨
沈航 陈娇 黎佰杨

摘要:为明确杂交水稻萌发率、苗期耐冷性与耐冷性指标的关系,以重庆师范大学10个自育杂交水稻组合和糯稻89-1为材料,测定种子的生理指标、低温萌发率及苗期低温处理恢复生长后的死苗率。不同杂交水稻组合的可溶性糖含量、维生素C含量、过氧化氢酶活性差异明显,种子自身所含这4种物质与水稻低温下的萌发率可能具有一定关系;种子萌发对温度变化较敏感,每个组合中温度为6 ℃处理条件下种子萌发率均高于11 ℃,初步推断临界点以下低温层积对水稻种子的萌发有积极作用;试验结果表明,不同低温条件处理下水稻苗期死亡率有明显差异,温度越低,水稻幼苗受害越大,复温后死亡率越高;聚类分析将杂交组合分为A、B、C 3类,A组、B组生理指标大部分都高于C组,低温萌发不同杂交水稻组合内变异系数较大。总的来说,水稻种子耐冷指标对萌发率可能具有一定作用,在苗期,水稻对低温较敏感,随着时间和温度增长受影响越大,根据耐冷指标、萌发率、死苗率对不同水稻杂交组合进行分类,研究结果可以为选育耐冷性水稻品种提供理论依据。
关键词:糯稻89-1;杂交水稻;幼苗;耐冷;低温胁迫、萌发率
水稻是我国最重要的粮食作物之一,与国家粮食安全密切相关[1]。水稻种植常受气温影响,一般每4~5年就发生1次较大规模的低温冷害[2]。据统计,全世界有24个国家、1 500万hm2以上的稻作面积时常受到低温威胁,使水稻严重减产,低温冷害俨然已成为全球性自然灾害[3-6]。日本最早于20世纪30年代开始水稻的耐冷性研究[7],至今已有70多年的历史,我国从20世纪70—80年代开始水稻耐冷性研究[8],如今,国内外对于水稻的耐冷性研究已取得较好的进展,在耐冷性基因定位克隆、耐冷生理生化和分子机制等方面都有很大的进展。
研究水稻耐冷性的目的在于培育出耐冷性强的水稻品种,以减轻低温冷害对水稻的伤害。水稻耐冷性研究虽然取得了很多的成果,但选育出的耐冷性强的水稻品种却较少[9]。糯稻89-1是耐冷性极强的材料,其芽期、幼苗期的耐冷性均达到一级标准[10]。糯稻89-1能通过腋芽休眠越过冬季,次年萌发再生,越冬再生季产量达6 291 kg/hm2,与正季相当[11],糯稻89-1是用于研究水稻耐低温的珍稀材料。本研究以糯稻89-1为亲本与多个品种(系)进行杂交,对不同杂交组合后代的低温萌发以及苗期的耐冷性进行分析,以期为耐冷性水稻材料的筛选提供理论依据。
1 材料与方法
1.1 试验材料
以重庆师范大学自育的10个杂交水稻组合、糯稻89-1为材料(表1)。试验在重庆师范大学水稻育种基地进行,于2017年3月播种,8月收集种子,保存3个月后进行生理指标的测定和不同温度下的萌发试验。所配组合中糯稻89-1为一级耐冷性品种。
1.2 试验方法
1.2.1 种子生理指标的测定 精选饱满的籽粒进行测定。各组合分别做3次重复,取平均值。可溶性糖(SS)含量的测定采用蒽酮-硫酸比色法[12];过氧化氢酶(CAT)活性的测定采用高锰酸钾滴定法[13];丙二醛(MDA)含量的测定采用硫代巴比妥酸(TBA)显色法[12];维生素C含量的测定采用钼蓝比色法[14]。
1.2.2 种子萌发率 选取饱满的籽粒经常规方法消毒后,置于培养皿中。先用蒸馏水侵泡10 min后,去除半饱满籽粒。用3%的次氯酸侵泡30 min后,用去离子水冲洗干净并均匀平铺于无菌培养皿中,放置于不同温度下培养,其間每天定时换水并保持培养皿湿润。设置的温度分别为6、11、16、27 ℃,12 d 后统计萌发率,各组合3次重复,取平均值。12 d 后,将6、11 ℃条件下未萌发的水稻种子转入27 ℃条件下继续萌发,12 d后统计萌发率。
1.2.3 苗期耐冷性鉴定 取不同杂交水稻组合种子100粒,灭菌后置于27 ℃条件下,在0.8%琼脂糖培养基上培养,待幼苗长至3叶1心时,留50株长势相近的幼苗移至温度为6、11、16 ℃条件下培养12 d后测量苗高、根长,统计死亡率,12 d后,将温度为6、11 ℃条件下的水稻幼苗移至27 ℃条件下恢复生长7 d,统计死苗率。3次重复,取平均值。
1.3 数据处理
采用Microsoft Excel 2010处理试验数据和绘制图表,用SPSS 22.0进行平均值、单因素方差、相关性及聚类分析,聚类选用Ward法分析。
2 结果与分析
2.1 不同杂交水稻组合生理指标比较
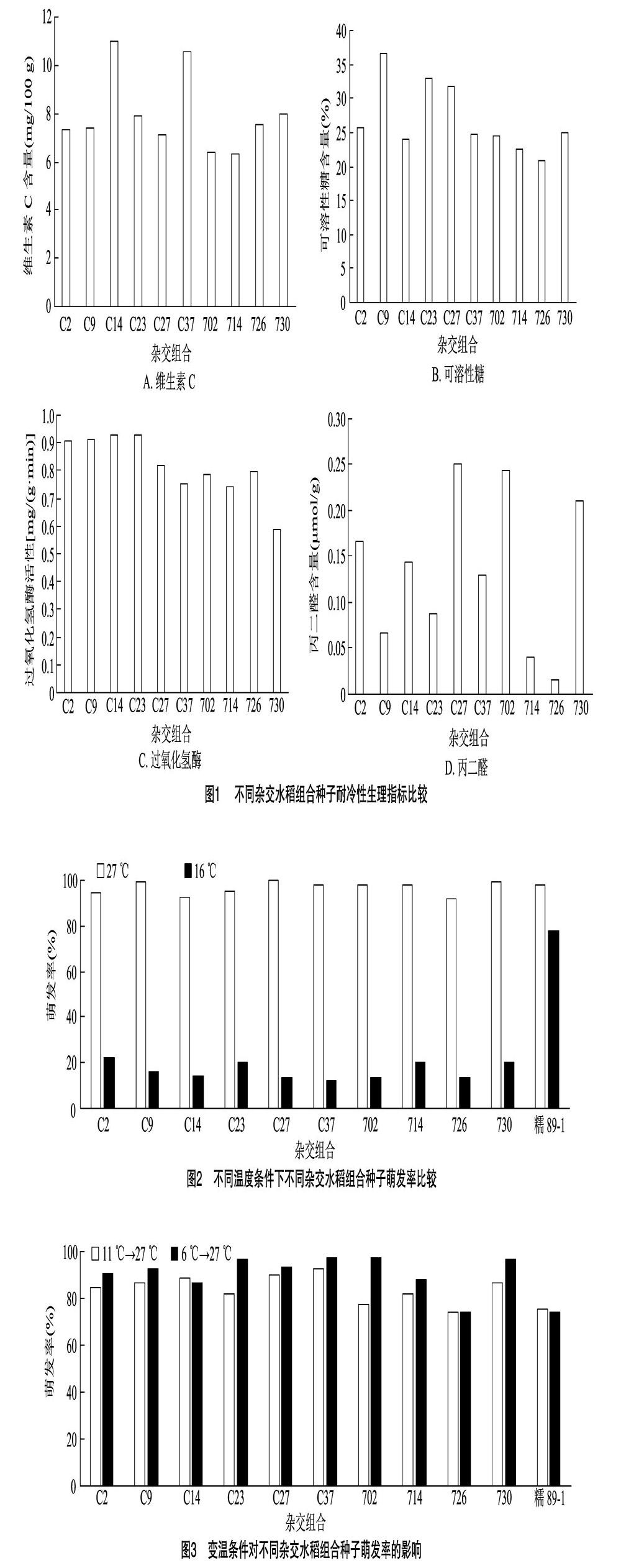
10个供试杂交水稻组合不同生理生化指标差异明显,MDA含量的差异性最大,CAT活性差异性最小。其中,维生素C含量变化范围为6.325~10.983 mg/100 g(图1-A);可溶性糖含量变化范围为20.73%~36.52%,平均值为26.84%(图1-B);过氧化氢酶活性变化范围为58.9~92.8 H2O2 mg/(g·min)(图1-C);丙二醛含量变化范围为0.016 4~0.250 4 μmol/g(图1-D)。726杂交水稻组合各项指标均低于平均值,MDA含量比平均值降低726.59%。
2.2 不同温度条件下不同杂交水稻组合种子萌发率比较
为明确种子耐冷性生理指标对萌发率的影响,对10个不同杂交水稻组合进行不同温度下种子萌发试验。恒温萌发条件下,温度为27 ℃时萌发率为96.67%,温度为16 ℃时种子虽能萌发,但萌发率均低于23%(图2)。变温萌发中,温度为6、11 ℃的条件下培养种子12 d后萌发率仍为0,继转入温度为27 ℃萌发,培养1 d后开始露白;6 ℃转至27 ℃的平均萌发率为91.29%,11 ℃转至27 ℃的平均萌发率为84.48%。每个组合中6 ℃处理过的种子萌发率均高于11 ℃。但6 ℃转至27 ℃萌发率低于27 ℃条件下的萌发率,可能是因为低温下处理的时间过长。不同低温处理对转温后不同杂交水稻组合种子萌发率的影响有所差异,其中对C23、702、730的杂交水稻组合影响最大,但对726杂交水稻组合影响较小。且726在不同温度条件下的萌发率均为最低,与耐冷性生理指标相对应(图3)。
糯稻89-1/CDR22与川农1A、Ⅱ-32A、宜香1A杂交组合归为A组,糯稻89-1/桂99与中9A、科2A、巴1A 3个不育系所配同父异母组合归为B组。4个不同恢复系与冈46A不育系所配同母异父杂交组合归为C组。A组、B组维生素C含量、可溶性糖含量、过氧化氢酶活性平均值均高于C组,温度为27 ℃条件下种子萌发率是低温(16℃)条件下的5倍以上,低温萌发不同组合内的变异系数较大。变温下,温度为6 ℃处理的种子萌发率均大于 11 ℃,表明不同的低温处理对种子萌发有明显的影响(表2)。
2.3 低温胁迫对水稻幼苗及恢复生长的影响
从图4可以看出,低温处理下,水稻幼苗普遍生长缓慢或接近停滞,并出现干枯腐烂的现象,且不同低温胁迫对幼苗的影响差异明显。温度为16 ℃条件下,不同水稻杂交组合幼苗均能够继续生长,但生长速度较27 ℃慢,培养3 d后,C9、C2杂交组合叶尖开始出现变黄迹象,继续培养 9 d 后发现不同水稻杂交组合叶尖均有不同程度的变黄,730、702、C37、C9杂交组合叶尖变黄程度严重,由叶尖变黄转为整叶变黄,最大变黄长度达3.3 cm,而糯 89-1 及726、714、C14杂交组合生长状态良好,叶尖变黄株数少。温度为6、11 ℃低温处理下,不同组合后代死苗率差异较大,糯89-1和714在2个温度条件下死苗率均为0,温度为11 ℃条件下幼苗死亡率普遍高于6 ℃ 条件下。27 ℃恢复生长7 d后,湿度为6 ℃处理的幼苗死亡率明显高于11 ℃处理的幼苗,且除糯89-1外的幼苗经过恢复生长期几乎全部死亡,C23、714杂交水稻组合由0上升至100%。C9、726、702杂交水稻组合幼苗死亡率相对较低,分别为84.00%、84.16%、77.86%。湿度为11 ℃处理下的幼苗死亡率虽然较恢复生长前有不同程度的增加,但大多数组合能恢复正常生长,其中C2、714杂交水稻组合幼苗死亡率较低,分别为4.05%、6.25%。
3 讨论与结论
植物种子播种后能否正常萌发是不同的内源性和环境因素相互作用,共同调节种子休眠与萌发之间的平衡的结果[9,15-16]。10个水稻杂交组合可溶性糖含量、维生素C含量、过氧化氢酶活性差异明显,这与不同水稻杂交组合遗传物质的差异有关。3个指标与种子低温下萌发率以及低温处理后恢复正常温度下的萌发率具有相关性,可能是种子可溶性糖含量、维生素C含量、过氧化氢酶活性与低温下种子的萌发率有关,本试验的结果与张启雷等的研究结论[17]相同。
种子萌发对温度的变化较敏感,低温是种子萌发的胁迫因素之一[18]。低温环境下,水稻种子萌发率会降低,其降低程度与种子耐低温能力有关。本研究中6、11 ℃低温条件下,不同水稻杂交组合萌发率均为0,说明该温度抑制了种子的萌发。温度为16 ℃时不同水稻杂交组合种子均有萌发,但萌发率普遍较低,糯89-1在16 ℃萌发12 d萌发率仍然能达到78%,说明低温耐性强,这与赵正武等的研究结果[18]具有一致性。本研究中,不同水稻杂交组合种子在不同温度处理后,恢复正常温度萌发时其萌发率均具有明显差异,初步推断种子萌发阶段,低温胁迫的强度对于胁迫后水稻种子活力的恢复有重要的影响。不同水稻杂交组合的种子在经过6、11 ℃ 低温处理后,其萌发率仍保持在74.07%~96.39%之间,大部分甚至高于优良亲本糯 89-1,其中涉及的潜在育种可利用潜力以及对深入研究种子经历低温后萌发的生理机制和遗传效应都具有重要的利用价值。当温度低于12 ℃时,遭遇中等倒春寒(日平均气温≤12.0 ℃ 4~6 d或日平均气温≤14.0 ℃ 7~10 d)甚至是强倒春寒(日平均气温≤12.0 ℃≥7 d或日平均气温≤14.0 ℃≥11 d)时,种子不能萌发,度过倒春寒后,气温回暖,种子活力基本恢复,方可继续萌发;除种子萌发受低温影响外,低温还会抑制幼苗的生长,使幼苗出现叶尖变黄、生长缓慢或存活率单向减少等现象[19-20]。低温引起植株叶片形态的变化是植物对冷害最直观的反映,主要包括叶片卷曲、萎缩、叶色褐化,甚至干枯死亡[21]。本研究中,温度在6 ℃条件下,幼苗几乎不生长,并且叶片失绿变黄干枯。转入27 ℃恢复生长时,在温度为6 ℃条件下胁迫12 d的幼苗几乎全部生长霉菌死亡,温度为11 ℃条件下大部分组合能度过恢复生长期,正常生长,低温胁迫严重影响幼苗生长,胁迫期间温度越低,对幼苗造成的伤害越大,在恢复生长期死亡率越高。此外,水稻杂交组合702尽管在冷害时出现整株枯黄的现象但在温度为27 ℃复育后死亡率最低,这可能与越冬再生的性状有关。
聚类分析的结果表明,10个水稻杂交组合的低温耐受性与糯89-1差异较大。水稻杂交组合C2、C9、C14以及C23、C27、C37为同父异母组合,714、726、730为同母异父组合,这9个组合并未按同父异母或是同母异父的组配方式聚为一类,各组合低温耐受性差异不大,说明苗期的耐冷性不存在细胞质遗传,而是受细胞核基因控制的,刘建丰等研究发现,杂交稻F1苗期耐冷性倾向于母本,受父本影响较小[22-23],出现不同结果可能是由于试验所利用的低温处理条件和耐冷性评价标准以及试验材料耐冷性程度不同。另外,除了702,其他9个组合均有糯89-1血缘,这9个杂交组合的苗期耐冷性与702相差不大,表明糯89-1耐冷性的遗传力较低,这与金润洲对耐冷性遗传研究的结果[24]相反,可能是所用材料的不同导致的。
水稻种子在低温胁迫下的萌发率很大程度上由自身生理物质含量的高低决定,而丙二醛含量、可溶性糖含量、維生素C含量、过氧化氢酶活性高低对水稻种子在低温胁迫下种子的萌发情况可能起到关键作用。水稻在苗期遇到冷害会影响秧苗的发育,不同杂交组合受到低温影响的表现基本相同,这与潘孝武等的试验结果[25]基本一致。本研究主要致力于挖掘以糯稻89-1为主要亲本的不同杂交水稻组合的耐冷生理指标与萌发率和苗期耐冷能力的关系,在涉及到糯稻89-1耐冷性状遗传方面还须进一步深入研究。
参考文献:
[1]邓兴旺,王海洋,唐晓艳,等. 杂交水稻育种将迎来新时代[J]. 中国科学(生命科学),2013,43(10):864-868.
[2]高益波,景元书,刘明璐,等. 抽穗扬花期低温强度对水稻生长发育的影响与模拟[J]. 江苏农业科学,2018,46(5):53-57.
[3]Sthapit B R,Witcombe J R. Inheritance of tolerance to chilling stress in rice during germination and plumule greening[J]. Crop Science,1998,38(3):660-665.
[4]王士强,赵海红,赵黎明,等. 水稻冷害生理功能变化与调控研究进展[J]. 中国农学通报,2017,33(36):1-6.
[5]王主玉,申双和. 水稻低温冷害研究进展[J]. 安徽农业科学,2010,38(22):11971-11973.
[6]崔 迪,杨春刚,汤翠凤,等. 低温胁迫下粳稻选育品种耐冷性状的鉴定评价[J]. 植物遗传资源学报,2012,13(5):739-747.
[7]Nishiyama I. Proceedings of the symposium on climate and rice[M]. Manila:IRRI,1976:159-185.
[8]韩龙植,张三元. 水稻耐冷性鉴定评价方法[J]. 植物遗传资源学报,2004,5(1):75-80.
[9]王尚明,贺浩华,肖叶青,等. 水稻东野1号苗期耐冷性遗传分析[J]. 湖北农业科学,2008,47(1):1-4.
[10]王春萍,雷开荣,李正国,等. 低温胁迫对水稻幼苗不同叶龄叶片叶绿素荧光特性的影响[J]. 植物资源与环境学报,2012,21(3):38-43.
[11]赵正武,李仕贵,雷树凡. 糯稻89-1越冬性遗传研究[J]. 中国农业科学,2006,39(12):2399-2405.
[12]赵世杰,史国安,董新纯. 植物生理学实验指导[M]. 北京:中国农业科学技术出版社,1998:136-139.
[13]汤绍虎,罗 充. 植物生理学实验教程[M]. 重庆:西南师范大学出版社,2012.
[14]孙 群,胡景江. 植物生理学研究技术[M]. 杨凌:西北农林科技大学出版社,2006.
[15]简水溶,万 勇,罗向东,等. 东乡野生稻苗期耐冷性的遗传分析[J]. 植物学报,2011,46(1):21-27.
[16]Shu K,Meng Y J,Shuai H W,et al. Dormancy and germination:how does the crop seed decide?[J]. Plant Biology,2015,17(6):1104-1112.
[17]张启雷,刘 强,高 辉,等. 内源抗坏血酸对水稻种子萌发及幼苗生长的影响[J]. 热带亚热带植物学报,2016,24(3):273-279.
[18]赵正武,王述民,李世平. 珍稀稻种资源越冬糯稻89-1研究初报[J]. 杂交水稻,2000,15(3):3.
[19]李太贵,王 磊. 低温下水稻幼苗形态生理应变研究[J]. 生态学杂志,1996,15(6):61-63,50.
[20]Mahajan S,Tuteja N. Cold,salinity and drought stresses:an overview[J]. Archives of Biochemistry and Biophysics,2005,444(2):139-158.
[21]许 英,陈建华,朱爱国,等. 低温胁迫下植物响应机理的研究进展[J]. 中国麻业科学,2015,37(1):40-49.
[22]刘建丰,陈立云. 水稻耐冷性研究现状与展望[J]. 作物研究,1996,10(2):42- 44.
[23]李 平,刘鸿先,王以柔,等. 低温对杂交水稻及其亲本三系始穗期旗叶光合作用的影响[J]. 植物学报,1990,32(6):456-464.
[24]金润洲. 日本关于水稻耐冷性鉴定及其遗传的研究[J]. 水稻文摘,1990,9(3):1-5.
[25]潘孝武,李小湘,黎用朝,等. 湖南早秈稻品种芽期和苗期耐冷性初步研究[J]. 杂交水稻,2015,30(5):59-64.邓惠清,唐灿明,刘正銮,等. 陆地棉空间诱变SP1~SP3主要性状变异分析[J]. 江苏农业科学,2020,48(12):64-67,74.
