微生物利用二氧化碳合成燃料及化学品
——第三代生物炼制
2020-07-15王凯刘子鹤陈必强王萌张洋毕浩然周雅莉霍奕影谭天伟
王凯,刘子鹤,陈必强,王萌,张洋,毕浩然,周雅莉,霍奕影,谭天伟
(北京化工大学生命科学与技术学院,北京 100029)
近年来,大量的化石燃料燃烧和过量的温室气体排放已严重影响环境和气候[1-2]。大气中的CO2浓度在过去的4万年中一直稳定在200~280μL/L[3],但在最近50年中,浓度急剧上升至近400μL/L,这种非线性增长仍在持续,到2050年,二氧化碳水平很可能会达到500μL/L[4],与1900年的水平相比,这可能将导致全球平均温度上升2°C。如此严重的全球温度变化将不可避免地增加冰川崩解的风险并造成负面的连锁效应[5]。随着合成生物学技术的不断发展,科学家渐渐开辟了一条为人类活动提供绿色环保替代品的道路(利用微生物生产生物燃料及化学品),以期达到节能减排的社会生产与发展模式。例如不断提升汽油中生物乙醇的混合比例,从而降低温室气体的排放并一定程度地减少对化石燃料的需求。
第三代生物炼制概念的提出[6],旨在利用大气中的CO2和可再生能源,例如光、废水中的无机化合物、光伏电池和风能等可持续资源产生的电能来进行生物生产(图1)。与第一代和第二代生物炼制相比,第三代生物炼制极大地降低了原料加工成本,对食品和水源供应的安全威胁降低很多。因此,研究人员在第三代生物炼制(CO2利用)方面已经取得了较大的进展,例如,已经验证了多种天然和合成的CO2固定途径,并建立光能电能的能量捕获技术,部分微生物固碳技术已成功应用并在商业化模式下进行运转,例如Lanza Tech 公司与宝钢集团合作建立的利用钢厂废气CO、CO2等气体进行生物乙醇的生产[7]。以CO2为原料的微生物制造的关键挑战是有效地固定大气中的二氧化碳和有效地捕获可再生能源用于生物生产。自养微生物可以利用CO2维持细胞生长,但是它们可能无法在工业条件下定向地高效地生产燃料或化学物质。为了实现CO2微生物利用的目标,已经利用合成生物学的基因编辑手段将自养生物进行改造以进行化学品的生产[8-10],另外,也将CO2固定途径整合到了异养微生物细胞工厂中进行CO2的利用[11-14]。
本文作者汇总了当前已验证的CO2固定途径的全面数据集,包括关键蛋白、酶动力学、产物以及能量消耗等;并分析了微生物CO2利用下的不同能量捕获技术,包括光自养合成和自养电合成,提出了与每种技术适配的菌株的选择方案;最后我们以对未来工程方向的讨论作为结束。
1 CO2微生物固定途径
在过去的40 亿年中,自然界已经发展出多样而复杂的二氧化碳固定途径。迄今为止,已经验证了几种固定二氧化碳的途径,并提出了不同的理论途径。在下文中,我们介绍了这些途径,并讨论了它们当前在能量供应、酶动力学、碳物种和浓缩机制等方面的局限性。
1.1 已验证的二氧化碳固定途径
迄今为止,已被验证的二氧化碳固定途径可以根据其特征(例如碳固定反应和固定的碳种类)分为六类,并在几篇文章中进行了一定的总结,而本文着重于阐述每种途径的共有与独特的特征(表1)[15-27]。
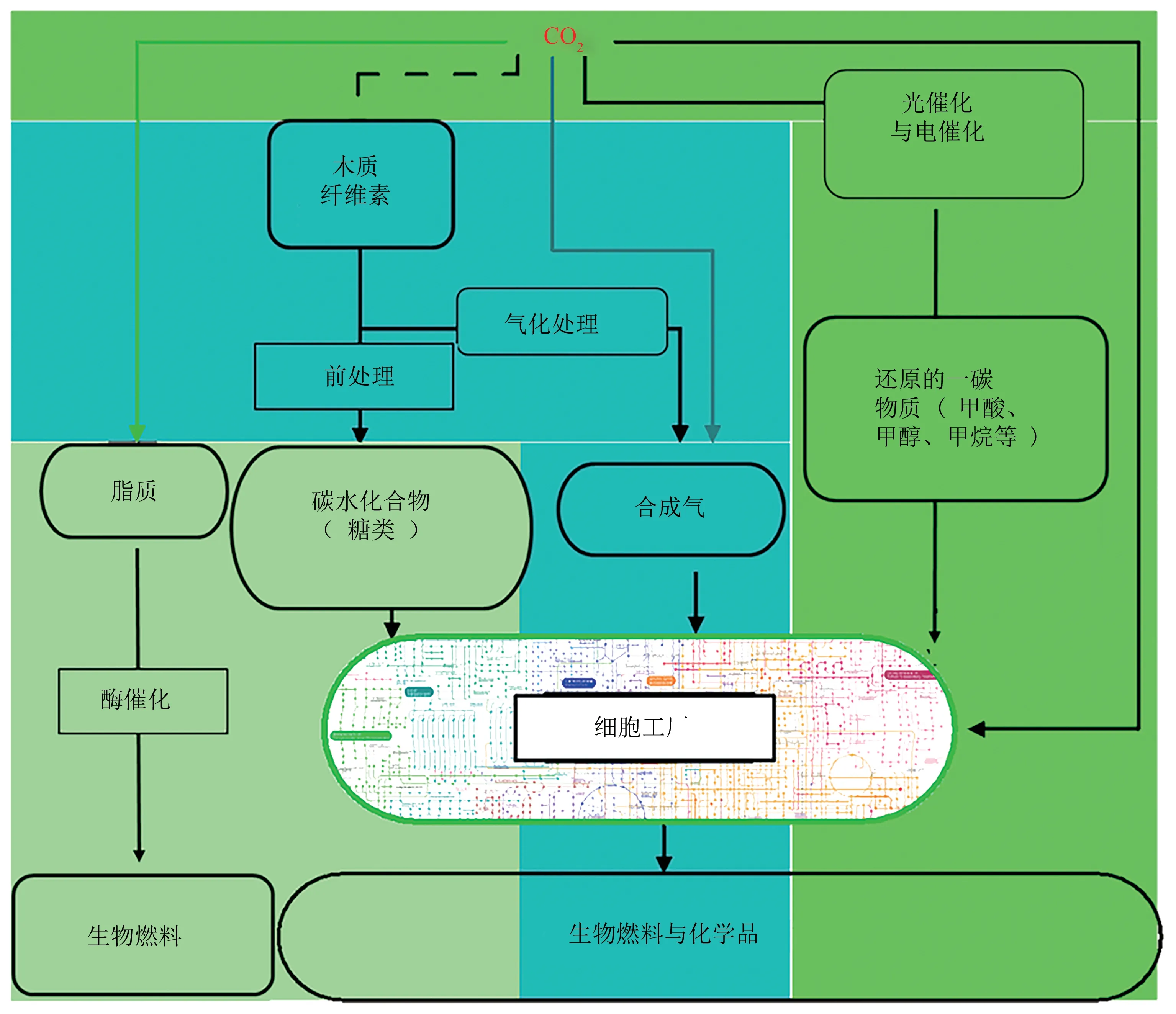
图1 三代生物炼制概述Fig.1 Overview of three generations of biorefineries
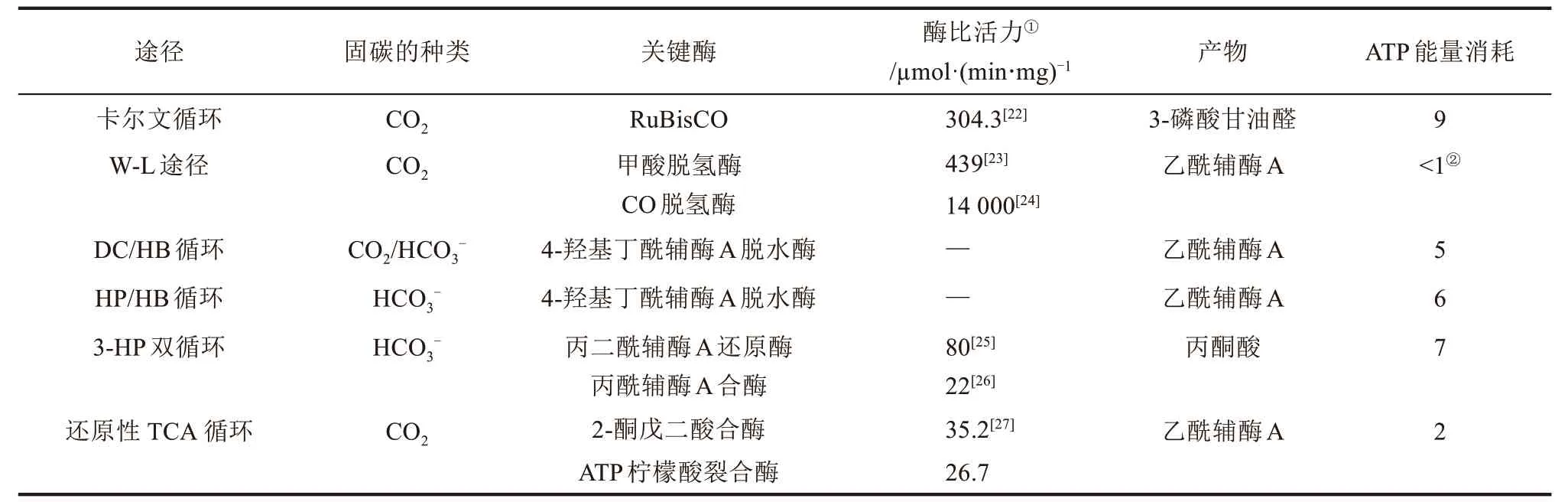
表1 天然二氧化碳固定途径的比较[21-27]Tab.1 Comparison of natural carbon dioxide fixation pathways[21-27]
Calvin-Benson-Bassham 循环(CBB 循环,也称卡尔文循环,或还原性戊糖磷酸循环)以碳水化合物为中心,并且与戊糖磷酸途径密切相关。大多数植物、藻类、蓝细菌和蛋白细菌都使用CBB 循环进行碳物质的利用[17]。CBB 循环中的关键酶是1,5-二磷酸羧化酶/加氧酶(RuBisCO)[18],而1,7-双磷酸七庚糖酶的过表达也增加了光合速率和细胞生长,表明该酶在CBB 循环通量控制中有着一定的作用[19]。最近,一个完整的CBB 循环被引入到大肠杆菌中,并以甲酸(也可以通过电化学方式由CO2生成的甲酸)作为电子供体,从而实现了大肠杆菌完全由CO2供能供碳的自养生长模式[20]。
Wood-Ljungdahl 途径(W-L 途径,也称还原性乙酰辅酶A 途径)[28]和还原性甘氨酸途径[29]是直接减少CO2的途径。这两种途径的拓扑结构非常相似。例如,首先将CO2还原并附着到C1载体上,然后再附着到另一个CO2分子上以生成C2化合物。Wood-Ljungdahl 途径中的关键酶是CO脱氢酶与甲酸脱氢酶[30]。Wood-Ljungdahl 途径在多种生物中具有活性,包括变形杆菌、扁平菌和螺旋线虫[30]。最近,Papoutsakis 教授团队在丙酮丁醇梭菌中表达了来自Clostridium ljungdahlii的Wood-Ljungdahl 途径的11 个核心基因,并报道了两个CO2固定分支都在丙酮丁醇梭菌中起作用。但是,连接两个分支的反应需要进一步优化[31-32]。在另一项研究中,研究人员发现还原性甘氨酸途径中的限速步骤是由还原甘氨酸裂解复合物催化的,还原性甘氨酸途径被认为是可行的二氧化碳固定合成途径[33]。最近,Figueroa 团队研究显示在磷酸盐氧化细菌假丝酵母厌氧菌中可能存在天然的还原性甘氨酸途径,但是仍然需要对该菌株进行彻底的生化分析,以确定还原性的甘氨酸途径是否自然存在并可以支持自养生长[34]。用于生产细胞甘氨酸和丝氨酸的还原性甘氨酸途径的异养表达已在大肠杆菌[29,35-36]和酿酒酵母[37]中得到证实。
二羧酸/4-羟基丁酸(DC/HB)循环[38]、3-羟基丙酸/4-羟基丁酸(HP/HB)循环[39]、3-羟基丙酸(3-HP)双循环[25,40]和还原性TCA 循环[41]是围绕常见的中间体演化而来的。例如,这些途径均利用两种保守的代谢产物琥珀酰辅酶A 和乙酰辅酶A,并且每个循环与该组中的另一个循环共享多个反应。根据固定的碳种类,这4个途径被进一步分为3类:
(1)DC/ HB 循环通过丙酮酸合酶固定1mol CO2,通过磷酸烯醇丙酮酸(PEP)羧化酶固定1mol碳酸氢盐。DC/HB循环中的关键酶是4-羟基丁酰辅酶A 脱水酶[42]。这种含FAD 的酶含有一个对氧不稳定的铁硫中心,但具有足够的耐氧性[42]。迄今为止,已经在厌氧菌中发现了这种循环,但尚未报道该循环的异源表达,该途径需要各种铁硫蛋白和硫酯[38]。
(2)HP/HB 循环[39]和3-HP 双循环[25,40]通过乙酰辅酶A/丙酰辅酶A羧化酶吸收2mol碳酸氢盐。这两个循环都具有非常高的能量需求,并且这些循环能够通过进化而留存下来的原因可能是由于它们可以耐受氧气并吸收碳酸氢盐(而不是二氧化碳)。吸收碳酸氢盐是有利的,因为细胞内碳酸氢盐的浓度可以比细胞内CO2的浓度高得多。HP/HB 循环中的关键酶是4-羟基丁酰辅酶A脱水酶[42],到目前为止,该循环仅在有氧古生菌株中发现[30]。最近,Keller 团队在热球菌中表达了来自景天金属小球藻的5 个HP/HB 周期基因,并成功地从H2和CO2中产生了3-羟基丙酸[43]。另一方面,3-HP双循环中的关键酶包括丙二酰辅酶A还原酶[44]和丙酰辅酶A合酶[45]。最近,Way等将3-HP双循环分为4个亚组,并在大肠杆菌中分别表达它们,这表明所有亚组都可以补充宿主突变,但是,所有3-HP双循环基因的异源表达未能使菌株进行自养生长[46]。为实现3-HP双循环的自养生长,可能需要减轻代谢途径中次级代谢产物对细胞生长的有害影响,可以通过电合成改善还原力的供给,并优化总体碳通量。
(3)还原性TCA 循环通过逆转氧化TCA 循环而固定了2mol CO2。还原性TCA循环中的关键酶包括ATP柠檬酸裂合酶和2-酮戊二酸合酶[21,42]。长期以来,人们认为柠檬酸合酶催化了乙酰辅酶A和草酰乙酸不可逆地形成柠檬酸。因此,为了自养生长,必须用ATP柠檬酸裂合酶或柠檬酸辅酶A合酶[47]与柠檬酸辅酶A裂合酶[48]代替柠檬酸盐合酶。最近的研究表明,在乙酸盐脱硫杆菌中[49],天然柠檬酸合酶可以催化正向和反向反应,但是,这些柠檬酸盐合酶是否能够支持重组宿主中的细胞生长尚不清楚。已经在绿硫细菌和水生细菌中发现还原性TCA 循环[30]。Liu等将还原性TCA循环整合到大肠杆菌的周质中,并使葡萄糖中苹果酸的产量翻了一倍[50]。
1.2 二氧化碳固定途径新思路
在现有的CO2固定途径外,研究人员对一些未知的固碳途径也进行了一定的探索。对许多自养生物的基因组分析显示,某些物种,如嗜酸亚铁杆状菌和砷热解热菌,不具有任何已知的CO2固定途径的基因[51],这表明可能还有其他尚未被鉴定的自养途径。潜在的CO2固定途径的鉴定需要对更多生物进行基因组测序,以确定它们是否使用已知的CO2固定途径,并进行详细的生化分析,然后重建和验证生物途径和网络。
另外,研究人员还提出了多条用于固定二氧化碳的理论途径。例如,已经提出的结合3-HP 双循环和DC/HB 循环中的反应步骤而形成的一种新的碳固定途径,其可通过丙酮酸合酶固定1mol CO2同时经由PEP 羧化酶固定1mol 碳酸氢盐,仅经过6 个步骤即生成乙醛酸[15]。Bar-Even 教授团队使用建模方法分析了5000 种代谢酶,并根据拓扑、ATP 效率、动力学和热力学可行性探索了可能的固碳替代途径[52]。基于此,他们提出了丙二酰CoA-草酰乙酸-乙醛酸(MOG)途径,通过PEP羧化酶固定2mol 碳酸氢盐生成乙醛酸酯。MOG途径借用了C4植物中自然进化的机制,其中碳首先被PEP 羧化酶固定以生成草酰乙酸,然后生成苹果酸,最后将苹果酸脱羧形成丙酮酸以完成循环。但在MOG 途径中,释放的CO2由PEP 羧化酶使用,而不是由RuBisCO 使用,就像在C4循环中一样。有人认为,就途径的特异性、动力学和ATP 效率而言,MOG 途径优于卡尔文循环[52]。
总之,对具有更高比活力并对碳酸氢根具有更高亲和力的羧化酶的探究,对于开发新的CO2固定途径起着一定的促进作用。
2 CO2固定途径中的关键影响因素
为了在CO2固定途径中获得高的固碳效率与生物生产,了解每个途径的机制至关重要,例如途径中的氧敏感性、能量需求、碳的种类和富集机制等。
2.1 氧敏感性
是否具有在氧气存在下运行的能力是区分CO2固定途径的一个重要的因素。CO2固定途径中的氧敏感酶包括CO脱氢酶/乙酰辅酶A合酶、丙酮酸合酶、铁氧还蛋白依赖性2-酮戊二酸合酶和一些金属依赖性甲酸盐脱氢酶[42]。
通常,Wood-Ljungdahl途径只能在严格的厌氧条件下运行,因为它使用了铁氧还蛋白和对氧非常敏感的CO 脱氢酶/乙酰辅酶A 合酶。DC/HB 循环和还原性TCA 循环具有对氧敏感的酶,但可以在厌氧和微需氧条件下运行。相比之下,卡尔文循环、还原甘氨酸途径、3-HP 双循环和HP/HB 循环都可以在完全有氧的条件下运行[21]。但是,氧的敏感性在各种途径和生物之间有很大的不同,从而使某些对氧敏感的酶或途径在有氧条件下也可以发挥催化作用。例如,C3植物中的卡尔文循环具有耐氧性,但是,RuBisCO 可能会竞争性地氧化1,5-二磷酸核酮糖,并使光合效率降低20%~50%[53]。
有氧自养生长可以实现多种产品的生物合成,但是,必须注意如何提高这种类型生物的生产量,因为在O2呼吸中需要大量的还原力才能产生ATP;另一方面,厌氧自养生物通常生长速度低,细胞密度低,或者它们合成的ATP 含量太少而无法产生能量密集的产品[54]。然而,利用有氧固氮途径和厌氧固氮途径的微生物生产都具有与工业相关的滴度和生产率[55]。
2.2 能量需求
所需的还原当量(仅基于起始化合物和终止化合物中的电子数计算)在所有CO2固定途径中都明显相同,而ATP的要求却有很大差异,ATP的变化范围从小于1mol 到9mol。这些不同的ATP 需求可以从三个方面来解释:①有氧或厌氧代谢,通常,在有氧条件下活跃的途径比在无氧条件下活跃的途径消耗更多的ATP,并且氧气的利用可以提供大量的ATP;②还原当量和电子给体,例如,铁氧还蛋白(E'0=-430 mV)提供的能量驱动力比NAD(P)H(-320 mV)高;因此,用两种铁氧还蛋白替代NAD(P)H 可 提 供约20kJ/mol 的额外能量驱动[56]。③类似地,由于H2S 的燃烧热(ΔHc=519 kJ/mol)比元素硫的燃烧热(293kJ/mol)高,因此该电子给体/受体对可提供226kJ/mol 的额外能量(以电子给体/受体对H2O/O2为参比)[57]。
2.3 碳的种类和富集机制
在CO2的微生物利用方面,其使用的碳物种主要包括CO2和碳酸氢盐。CO2在与空气(pH 7.4,20℃) 平衡时,其在水中溶解的浓度仅为0.012mmol/L[42],由于该浓度高度依赖于温度和盐度[58],并且大多数生物体对高温和盐度敏感,因此很难在体内优化该值。另外,与空气(pH 7.4,20℃)平衡时的碳酸氢盐浓度为0.26mmol/L[42]。该值主要取决于溶解的CO2浓度和pH(pKa[HCO3-/CO2]= 6.3),在海水(pH7.8~8.2)中甚至更高。因此,使用碳酸氢盐的碳固定反应可能比使用二氧化碳的碳固定反应更有效。
底物浓度的增加可以改善热力学和酶转化效率,以及降低酶的副反应效率。可以通过依赖于能量的CO2捕获机制来优化CO2的浓度,包括使用CO2浓缩机制(CCM)、跨膜碳酸氢盐泵、转运蛋白[59]以及碳酸酐酶和羧化酶含量高的固碳细胞器微室等[60]。
3 CO2固定能量利用
碳从CO2(氧化态+4)同化为生物质(氧化态约0)需要大量能量,随着生物技术的发展以及对菌株的不断探索,现在微生物也可以从光能或电能收集中获取能量以支持其生长与生产,也称为人工碳固定系统(图2)。当前,微生物CO2的利用在碳利用效率方面落后于传统的五碳糖与六碳糖的利用效率。然而,其能量转化效率较高,例如,在五碳糖与六碳糖的利用中,太阳能到生物质到产品的总能量转换效率估计仅为约0.2%[61],而据报道光合自养生物的太阳能到产品的效率为1%~3%[61],化能自养微生物生成化学品(如H2)的效率约为7%[61],自养电合成微生物的太阳能到产品的生成效率可高达9%~10%[62]。
3.1 光能利用:光合自养
光合自养利用光子的能量将CO2转化为有机化合物。光合生物可分为产氧光合生物(如植物、藻类、蓝细菌)以及不产氧光合生物(如绿硫细菌)。产氧光合生物主要利用CBB循环,而不产氧光合生物利用各种不同的途径,例如CBB 循环与还原性TCA 循环[30]。最近的研究表明表达蛋白视紫红质光系统的大肠杆菌[63]和与光捕获纳米颗粒整合在一起的酿酒酵母[64]使用光生电子进行细胞生长和生产,为工业模式菌株的光合自养合成铺平了道路。但是,这些光合生物杂交系统仍处于开发的初期,其他挑战包括材料的生物相容性、光收集装置的选择以及生物和非生物成分的无缝耦联等方面仍需要进行不断的研究与探索。近期杨培东教授团队在光合生物杂交系统方面,作了较为全面的介绍[1]。
3.2 电能利用:电能耦合的生物合成
电能耦合的生物合成需利用电力,该电力可以通过多种可再生资源(包括光、风、潮汐、水力和地热)产生,将CO2转化为微生物系统中的燃料和化学物质。目前,CBB 循环[65]与Wood-Ljungdahl途径[66]已在电化学微生物体系中进行了一定的应用。
根据能量输送策略,电能微生物耦合系统可分为直接电荷转移系统和能量载体转移系统,微生物既可以直接吸收这些能量将CO2转化为有机化合物,也可以凭借能量载体转移系统来利用外源的电能,从而进行CO2的活化与利用[67]。一方面,外生电物种,例如梭菌和穆尔氏菌,可用于低驱动电压直接电荷转移系统,并表现出独特而有效的机制,促进电子在细胞膜和导电表面之间转移[68]。另一方面,在能量载体转移系统中,低驱动电压可用于产生能量载体,例如甲酸盐、氢、一氧化碳、甲醇、甲烷、氨、硫化氢和亚铁盐,以支持细胞生长[69]。H2、CO 和甲酸是厌氧条件下自养电合成的有吸引力的能量载体[69],但是由于H2和CO 是易燃气体,因此在有氧自养性电合成下将其用作电子载体可能会引发安全隐患。因此,我们认为甲酸盐在有氧条件下可能代表更有希望的能量载体,因为甲酸盐具有高溶解度和高氧化还原电位,但不需要额外的电子受体,也不会产生与挥发性有关的安全隐患。另一方面,由于甲醇具有与甲酸类似的高溶解度与利用效率,也被认为是一种良好的C1利用原料[35,70-72]。同样,近期的一篇综述文章在微生物的电能利用方面,对CO2固定、氮气还原和氨气生产进行了阐述,并且在电子转移机理、电极对生物电催化的影响、生物电催化生产生物燃料等方面进行了较为详细的总结[73]。

图2 人工碳固定系统Fig.2 Artificial carbon fixation system
4 展 望
微生物的CO2利用提供了通过闭环循环资源和减少二氧化碳排放以解决生态和社会问题的机会[74]。气候变化增加了人们对使用替代技术来生产燃料和化学品的需求的认识,而微生物的CO2利用提供了收集和循环利用CO2的机会。但是,考虑到实现该目标所需的高成本和大量时间的投资,仍然需要进一步增加社会、政治和经济激励措施。
如何确定理想的生产宿主也是一个较为困难的问题,但应考虑以下特征:原料耐受性(烟气或废物流),培养条件(开放池塘或密闭条件,淡水、废水或海水,氮源及能源等),目标产物(氧化或还原形式,有价值的产物或大宗化学品以及产物耐受性),能量捕获效率,碳固定效率,细胞生长率,生产能力(实际产量和生产效率),对污染和环境挑战的鲁棒性,细胞代谢过程,基因操作的可行性和稳定性。与特征明确的模型生物相比,工程自养生物的挑战可能包括其生长相对缓慢、培养策略复杂或缺乏有效的基因编辑工具;而将自养途径整合到模型异养生物中的挑战可能包括外源自养能量系统与宿主能量系统不适配、外源酶在宿主体内表达情况较差等[75]。有氧自养生物可能比厌氧自养生物更适合用于合成需要ATP的产物,而厌氧自养生物可能比有氧自养生物更适合自养电合成。此外,按比例放大仍然具有挑战性,因为为光自养生物提供足够的光、为有氧化学自养生物提供潜在的爆炸性气体混合物(O2、H2、CO 等)以及自养电合成的电子转移效率有关的困难仍然存在。在这里,我们建议采用有吸引力的宿主生物包括光合自养生物, 例如Scenedesmus obliquus(已在商业流程中使用[76])、梭状芽孢杆菌(可从烟道中去除95.9%的CO2、100%的SO2和84.2%的NO[77])、可实现高碳回收率的厌氧梭菌( 如Clostridium ljungdahlii、Clostridium autoethanogenum),以及大肠杆菌和酿酒酵母等模式生物。
总而言之,我们相信,尽管面临技术挑战和市场准入障碍,但随着近期技术的进步,微生物的CO2利用可能会对建立可持续发展的社会做出重大贡献。未来的研究方向应将微生物的CO2利用作为一个单独的研究模块,包括原料供应和耐受性、碳固定和利用技术、能量收集技术以及发酵工程技术等。
