滇东南喀斯特小生境土壤水来源与运移的稳定同位素分析
2020-07-15户红红
牟 洋, 范 弢, 户红红
(云南师范大学旅游与地理科学学院/云南省高原地理过程与环境变化重点实验室,云南 昆明 650500)
研究土壤水分运移对实现土壤水和地下水的有效利用具有理论和现实意义.稳定同位素作为理想示踪剂较早被应用于土壤水分运移的研究[1],并在土壤水—地下水—植物水—大气水等水分相互转化关系的研究中发挥着作用,蒸发引起的分馏效应使同位素值产生偏差和降水的差异性补给则是研究基础[2].国外较早根据稳定同位素的季节变化来研究不同剖面同位素组成、土壤水分入渗、径流和地下水补给量等[3-5],并建立相关模型.
喀斯特地区土壤水运移主要有优先流、活塞流两种方式.夏季降水很快通过大孔隙渗透,避免了蒸发损失,在土壤水和地下水补给中起主导作用,根区优先流活跃,而活塞流运移广泛存在[6-8].刘伟等[9]运用稳定同位素对比了喀斯特与非喀斯特地区土壤水运移特征差异,喀斯特地区土壤基质流较弱,而非喀斯特地区随土层深度增加,基质流混合作用增强.通过变异系数和标准差对比分析桂西北峰丛洼地典型坡面不同覆被及坡位的土壤水分动态变化及其空间变异,表明土壤水分空间差异及变异程度主要受前期降水量、有机碳、取样时间、地势和裸岩率的影响[10-11];表层土壤水稳定同位素变化较大,随土层深度增加变化减少,土壤水分对降雨响应的滞后时间与前期土壤含水率有关[12].对喀斯特峡谷地区不同生境的土壤物理特征和水分动态研究表明,各生境水分胁迫频繁发生,但出现的次数、程度、持续时间各不相同,其中干旱期失水最快的是石槽,其次是石沟,最慢的是土面[13].
滇中岩溶高原乡土树种滇青冈(Cyclobalanopsisglaucoides)及其伴生乔木水分利用层位呈现时空差异,旱季主要利用深层土壤水,并对表层岩溶带水利用增加,趋向于利用稳定的水源;雨季主要利用浅层土壤水[14].植物根系水力提升会对水分进行再分配,进而对土壤水分进行调节,但高粘土含量可能阻碍植物对土壤水提取[15].滇东南文山州石漠化面积广,占全省石漠化面积的27.5%,潜在石漠化土地面积大,作为石漠化恢复树种的云南松(Pinusyunnanensis)对贫瘠的土壤环境具有很强的适应能力,是优良的造林树种[16-17].石漠化程度不同,植物水分利用来源也存在差异,轻、中度石漠化环境下,植物主要利用土壤水;重度石漠化环境下,植物对降水的利用比例增大[18],这都会对土壤水的运移产生影响.
目前对普者黑峰林湖盆区内土壤理化性质的空间异质性及植物水分利用来源已有研究,但缺乏小生境尺度上土壤水分运移的影响研究,本研究通过分析3种小生境垂直剖面土壤水δ18O同位素的组成差异,探究小生境差异及其发育植被是否对土壤水运移产生影响,并为西南岩溶区现有植被恢复模式的合理性以及对喀斯特山地环境的适应性和指导农业生产提供依据.
1 研究区概况与研究方法
1.1 研究区概况
研究区位于云南省文山自治州丘北县普者黑(103°34′—104°45′E、23°45′—24°28′N),地处云贵高原南段和广西丘陵交界地带,石漠化广布.面积约165 km2,属中亚热带高原季风气候,年平均气温16 ℃,年平均降雨量1 214 mm,分为明显的干湿两季,雨季5—10月,旱季11月—次年4月.研究区内土壤类型为各类石灰土,土层厚度80 cm左右,不同生境有差异.受人类活动影响,原生林基本已被破坏,主要植被群落为清香木次生林、小叶羊蹄甲灌丛,以及后期种植的云南松人工林等,小生境类型差异明显,植被特征等见表1.植被—土壤层—表层裂隙共同构成该区域的表层岩溶生态系统,其对水分的蓄持为地下水提供水源,表层岩溶泉发育,是该地生态、生产和生活的稳定水源.

表1 不同群落小生境组合类型及特征
1.2 研究方法
1.2.1 生境调查 选取菜花箐山坡上坡位石沟、中坡位土面和下坡位土坡3个典型小生境进行调查取样,调查生境内植物种类、基岩裸露率、植被盖度、土壤类型、枯落物存量等.石沟为构造裂隙控制形态,其间土壤(灰棕色石灰土)充填[19],土石比≥70%,植物主要是萌生的矮小的小叶羊蹄甲(Bauhiniabrachycarpa),平均胸径1.7 cm,高度1.6 m,种植密度0.8株·m-2,基岩裸露率高,乔木层盖度低;土面发育在直径≥0.3 m圆状或近圆状的溶痕上,枯落物较多,土层较厚,为暗棕色石灰土,土石比≥40%,树种清香木(Pistaciaweinmannifolia),平均胸径13.4 cm,高度9 m,种植密度0.5株·m-2,乔木层盖度大;土坡枯落物存量高,土层厚,为黄棕色石灰土,土石比≥80%,树种为细叶云南松(Pinusyunnanensisvar.tenuifoli),平均胸径17.5 cm,高度12 m,种植密度0.3株·m-2,基岩裸露率低.
1.2.2 样品采集 选用研究区2014年5月—2015年4月自动监测站相关气象数据资料,主要有各月平均气温、月降水量等.
2014年5月—2015年4月一个水文年内,在每个月的一次有效降水后采集大气降水作为降水样,在露天空旷位置高处放置自制的采集器收集雨水.采集菜花箐泉作为地下水样,迅速用100 mL聚乙烯塑料瓶密封低温保存.
根系和木质部样品采集均在晴天上午,同一植株各采集两个平行样.采集清香木和云南松根系,在冠幅内中间位置按照10 cm递进挖取30 cm×30 cm×10 cm的土壤柱至1 m,用土壤筛手工选出根系,按根系直径<5 mm、5~10 mm、>10 mm分级,洗净后烘干至恒重计算根系干重.小枝采样在无人干扰或干扰较小的样点逐月进行,采集小叶羊蹄甲、清香木和云南松阳面栓化无叶片的小枝,长约3 cm,去除韧皮部保留木质部,迅速装入塑料离心管,密封保存.
雨后,在所选植物冠幅内的树根附近逐月对3种小生境用土钻法进行土样采集,取样深度为20、40、60、80 cm,分别取土样30 g,混合完全后放入样品袋密封.
1.2.3 样品测定分析 土壤性质测定:土壤容重、总孔隙度和毛管孔隙度采用环刀法,土壤含水率测定使用烘干法;土壤有机质测定采用重铬酸钾氧化—外加热法,土壤机械组成测定采用比重计法[20].
水样测定:土壤水和植物水样品采用真空抽提法获取[21].水样在中国地质科学院桂林岩溶地质研究所岩溶地质与资源环境监督检测中心采用Finnigan MAT-253质谱仪测定,测定结果用标准平均大洋水(V-SMOW)标准校正.δD和δ18O实验误差分别为0.5‰、0.1‰.
1.3 数据处理及制图
本研究运用Iso Source模型计算植物对潜在水源的利用比例,运行模型前需设定两个参数:一是来源增量参数设置为1%,表示以1%增量赋值植物对各潜在水源的利用比例;二是质量平衡公差设置为0.1%,表示各水源同位素值被利用比例加权值之和与植物木质部水同位素值的差异不超过0.1%时,该比例组合被认为是可能的组合[22-23].
采用Excel 2016对降水、土壤水和表层岩溶水样稳定同位素进行相关性分析和变异系数计算,并进行绘图.
2 结果与分析
2.1 不同小生境水体同位素组成特征及潜在水分来源
研究区降水水样δD和δ18O值的范围分别为-103.2‰~11.0‰、-14.4‰~0.4‰,表现出随降水量的增大而显著贫化的降雨量效应,但存在滞后效应(图1).研究区温度效应不明显,这与年温差较小,只在冬季温度偏低,而分为明显的干湿两季等因素有关.在5月(旱季末),稳定同位素达到峰值,随雨季到来降水增多,同位素逐渐贫化,8月降到最低值,西南季风携带水汽对研究区降水影响强烈.雨季结束后降水减少,同位素值逐渐升高,12月降水最少,稳定同位素富集明显,出现第二个峰值,后略有下降又不断升高.旱季降水同位素的高值说明夏季风输送水汽对研究区降水影响小,12月—次年2月降水稳定同位素值再次降低,过量氘增大,说明研究区局地水汽再循环强烈,对大气降水有显著贡献.
研究区降水线(local meteoric water line, LMWL)为δD=7.780 3δ18O+10.622 9(R2=0.97,n=12)(图2),与全球降水线(δD=8δ18O+10)相比斜率略小,截距偏大.斜率小表示大气干燥,分子质量小的氢同位素比氧同位素分馏速度快,δD偏重的程度大于δ18O,表现为普者黑地区截距偏大.
对小生境土壤水δD和δ18O拟合分析,石沟:δD=6.774 4δ18O-11.152(R2=0.952 2,n=48),土面:δD=7.573 4δ18O-1.485 6(R2=0.971 1,n=48),土坡:δD=7.082 3δ18O-10.477(R2=0.963 8,n=44).各小生境土壤水稳定同位素方程斜率和截距均小于研究区降水线方程,土面土壤水稳定同位素方程截距较石沟和土坡偏大,土壤水δD介于-125.8‰~-27.5‰,δ18O介于-17.4‰~-3.7‰,同位素组成主要落在当地大气降水线右下侧,表明大气降水是小生境土壤水主要补给源,生境不同蒸发强度不同,土壤水氢氧同位素发生不同程度的不平衡分馏,土坡土壤水稳定同位素较石沟和土面较偏负.各生境土壤水δD和δ18O分布集中,可能是降雨入渗与土壤中老水发生混合.研究区表层岩溶水δD和δ18O的变化范围分别为-75.3‰~-71.2‰、-10.7‰~-9.7‰,稳定同位素值落在降水线右下方,与降水稳定同位素相比较贫化,与土壤水相比较富集,对小生境土壤水进行了补给,其变化范围小于大气降水和土壤水,进而对土壤水氢氧同位素值产生影响.综上,降水和表层岩溶水为研究区土壤水主要的环境水源.
2.2 不同小生境土壤水δ18O变化特征
雨季,石沟和土面土壤水δ18O波动变化明显,土坡则呈现出快速贫化趋势,各生境表层土壤水δ18O值偏高于深层,土坡较石沟和土面土壤水δ18O偏负(表2),土壤含水率随深度逐渐增加,石沟和土面均呈波动变化趋势(表3).旱季,石沟土壤水δ18O先富集后贫化,土坡土壤水δ18O变化趋势与之相反,土面迅速贫化.各生境表层土壤水δ18O值均偏低于深层,土坡深层土壤水δ18O变化范围最小(-13.3‰~-5.3‰),土壤含水率随深度增加,土面与之相反,石沟土壤含水率低.
石沟雨季表层土壤水δ18O变异系数绝对值较下层大,旱季时表层和深层土壤水δ18O变异程度较低(0.16),土壤基质流混合作用较强,下层出现极大值(0.41).土面雨季各层土壤水变异程度均为各生境中最大,土壤水基质流混合作用较弱,旱季时变异程度较低,深层出现最低值(0.10).土坡雨季土壤水δ18O变异系数绝对值在40 cm层出现极值(0.38),表层变异系数大于深层,旱季时深层土壤水δ18O变异系数绝对值为各生境最大(0.27),其它各层均较小,土壤基质流混合作用强.
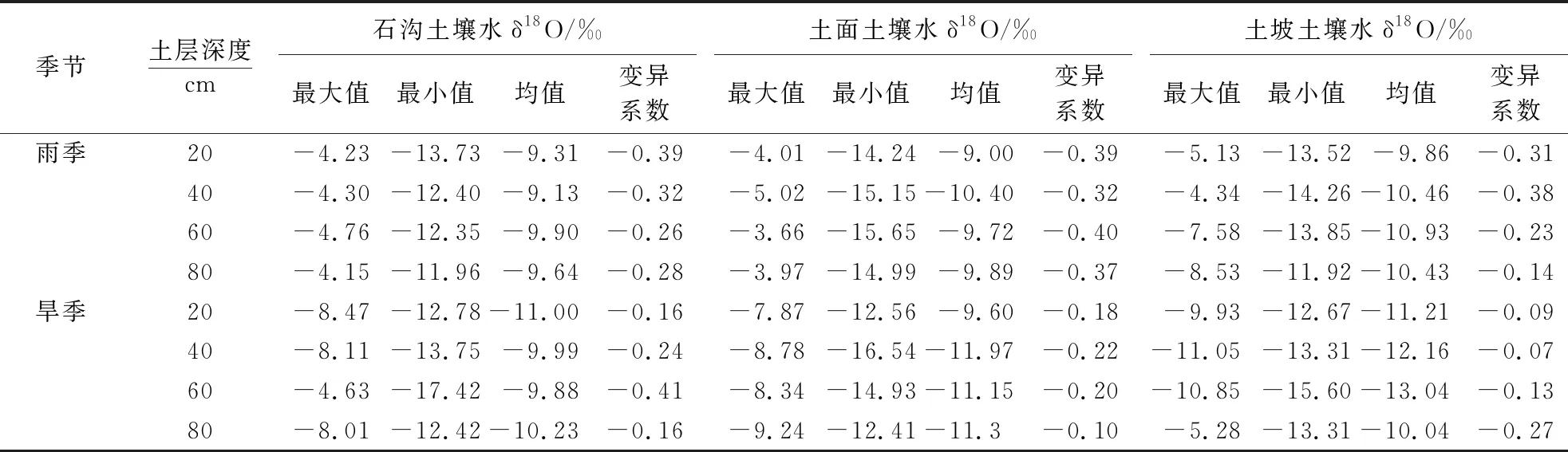
表2 不同小生境雨季和旱季土壤水δ18O统计特征1)
1)0~20 cm:表层,20~40 cm:中层,40~60 cm:下层,60~80 cm:深层.
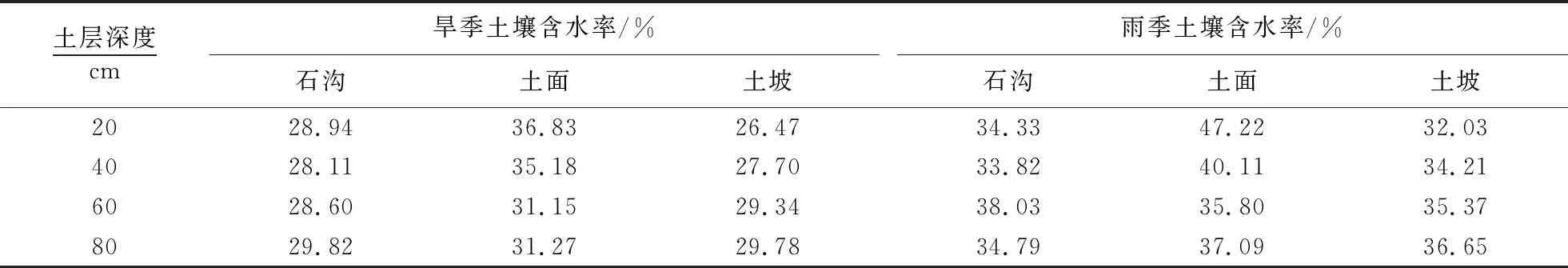
表3 不同小生境雨季和旱季土壤含水率1)
1)0~20 cm:表层,20~40 cm:中层,40~60 cm:下层,60~80 cm:深层.
2.3 不同小生境土壤水δ18O差异分析
2.3.1 石沟土壤水δ18O变化 土面和土坡表层为沙壤土,3种小生境表层以下土层均为黏土(表4),土壤基质流混合作用较弱,但均可能存在活塞流运移机制.石沟小生境雨季土壤水δ18O总体表现为逐渐偏负,60 cm层土壤含水率快速升高,大孔隙优先流明显.除5月外,各层土壤水δ18O变幅不大,随雨季到来土壤水δ18O值逐渐降低(图3).5月表层土壤水δ18O富集,下层迅速降低,其它月份石沟小生境土壤水δ18O随土壤深度呈波动变化,土壤水饱和,降水开始以活塞流入渗.
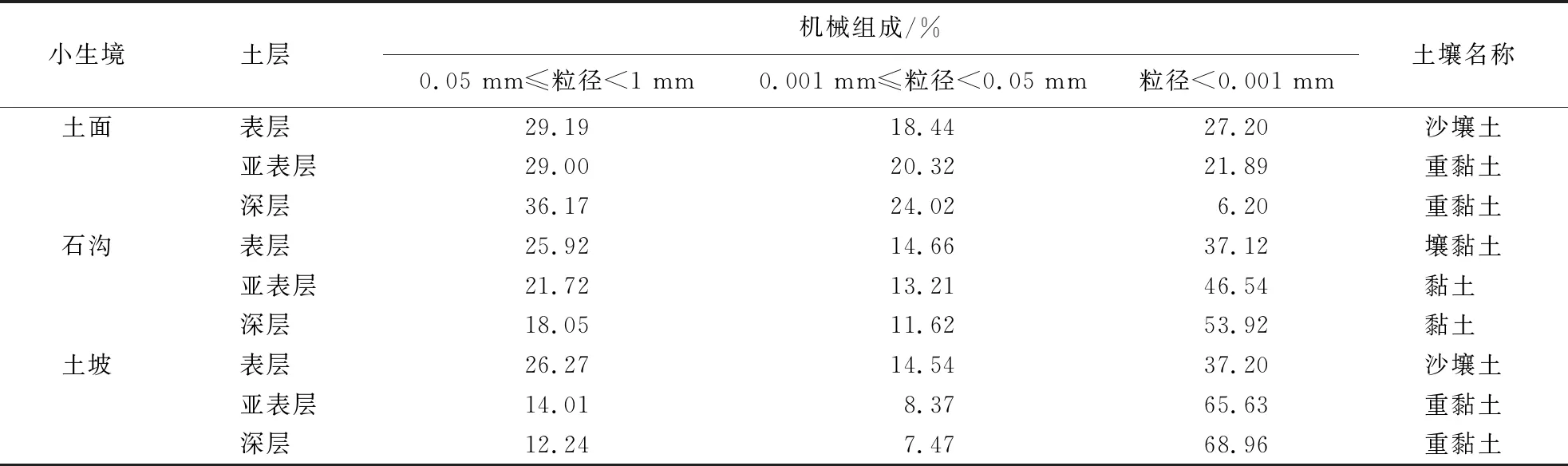
表4 不同小生境土壤机械组成1)
1)5~25 cm:表层,25~40 cm:亚表层,80~100 cm:深层,各生境有差异.
旱季,表层土壤水δ18O集聚,至60 cm层波动增大后逐渐收缩,是与石沟上层孔隙度大、容重小(表5),蒸发旺盛,60 cm层因大孔隙存在,土壤含水率升高,地下水对深层土壤水进行补给有关.雨季到来前表层土壤水δ18O富集,降水通过优先流形式迅速补给下层土壤水,使δ18O值降低.60 cm层土壤水δ18O波动强烈,变异程度高,表明土壤基质流混合作用微弱.1月上层土壤水δ18O几乎无变化,至60 cm层土壤水δ18O值达到最高后降低,这与旱季降水稀少,土壤水δ18O富集有关.4月60 cm层土壤水δ18O迅速降到最低值是因降水以优先流形式迅速补给该层土壤水,δ18O值迅速贫化[24].
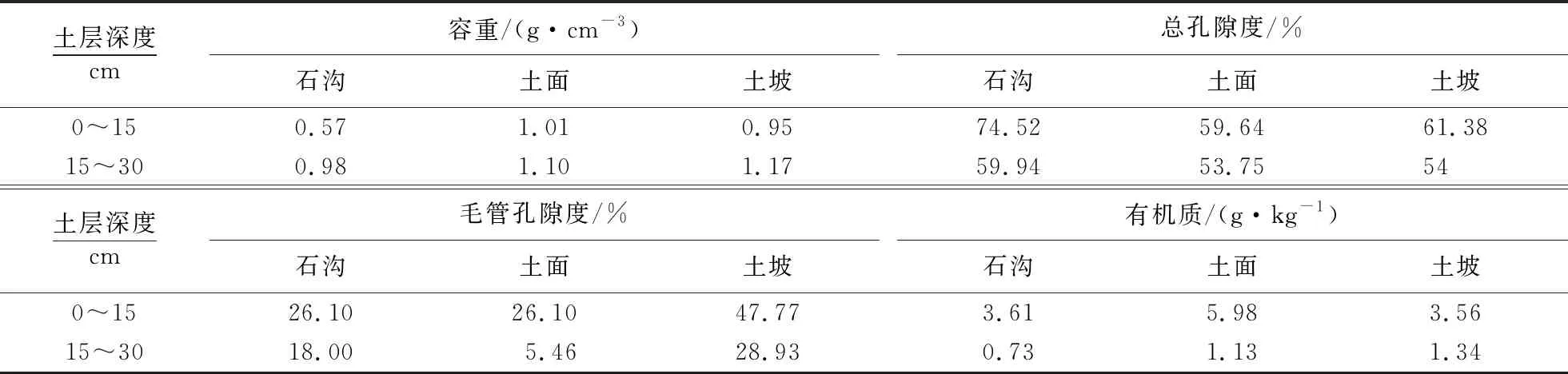
表5 不同小生境土壤理化指标
2.3.2 土面土壤水δ18O变化 土面小生境雨季和旱季土壤水δ18O随深度波动变化明显(图4),土壤水以活塞流入渗为主.因上层土壤非毛管孔隙度大,随雨季到来,降水快速下渗,40 cm层土壤水δ18O值迅速降低.6月土壤水δ18O富集,降水补给存在明显的滞后效应,9月土壤水δ18O才降到最低值,雨季土壤水δ18O总体表现为逐渐偏负.旱季土壤水δ18O变异较复杂,除1月外,40 cm层土壤水δ18O值均快速减小,可见蒸发对小生境表层土壤水δ18O存在影响,1月是旱季40 cm层土壤水δ18O唯一在该层位值升高的月份,土壤水δ18O富集.因前期土壤水滞留,3月植物根系水力提升作用使表层土壤水δ18O富集,下层迅速下降至不变.
2.3.3 土坡土壤水δ18O变化 土坡小生境雨季初和雨季末表层土壤水δ18O值较高,土壤水δ18O值变幅较大(图5),分别在60和40 cm层迅速降低,各月土壤水δ18O呈逐渐收缩趋势,原因是土坡上层土壤存在大孔隙,土壤水以优先流形式运移直接补给下层土壤,雨季初云南松对深层土壤水和表层岩溶水利用比例较高(表6),根系水力提升作用将下层土壤水提至上层,使浅层土壤水δ18O偏负;雨季结束,表层土壤水δ18O开始逐渐富集.旱季各月土壤水δ18O曲线呈波动变化,深层均逐渐富集,降水以活塞流方式补给土壤水.20~60 cm土壤水δ18O值贫化,变化幅度小,深层土壤水δ18O值差异较大.

表6 清香木和云南松根系分布特征
2.4 不同小生境植物水分利用分析
从表6可以发现清香木和云南松细根含量相当,云南松粗根含量高于清香木,并且两个树种的根系剖面分布类似,但是水分利用差别较大.根据Iso Source模型计算出植物对潜在水源的利用比例,可以发现石沟生境小叶羊蹄甲全年对各层土壤水和表层岩溶水均有利用(图6),水分利用策略属于机会型,5月对表层土壤水利用率达40.5%,8月对中层土壤水和表层岩溶水利用比例分别为27.7%、38.1%,10月对中、下层土壤水利用比例较高,其它月份对各层土壤水利用比较接近.雨季,土面生境清香木对表层岩溶水利用率高,在6月和8月对表层岩溶水的利用比例达到64.3%和78.8%,7月清香木可能是利用了前期降水,9月中、下层土壤水的利用比分别为32.1%、39.6%.旱季,1月清香木对中层土壤水利用比例高达72.8%,3月对上、中、下层土壤水利用比例分别为30%、27.3%和30.7%.6月时,土坡生境云南松对深层土壤水和表层岩溶水利用比分别为29%和35.2%,8月和9月时对岩溶水利用比高达93.4%和84.4%,10月对各层土壤水利用比例较为接近.旱季,12月和2月对下层土壤水利用比例为68.2%和34.8%,在4月对表层土壤水利用比例达到52.8%,对下层的土壤水利用比例只有6.6%,其它月份可能利用了前期降水.正是因各生境上发育植被的异同,对土壤水的利用策略差异,从而可能会对各生境土壤水的运移产生影响.
3 讨论
3.1 喀斯特土壤水运移机制
喀斯特土壤水运移方式主要有向下补给的活塞流、优先流和向上的地表蒸发、根系水力提升两种.土壤水δ18O在垂直方向存在波动变化,是因降水入渗后与土壤水混合后以均质活塞流形式层层向下推进引起的[25].当某一层位土壤水δ18O值迅速降低,就可能是降水通过大孔隙以优先流的形式迅速补给土壤水,使土壤水δ18O值降低.研究区土壤水δ18O主要落在当地大气降水线右侧,部分落在大气降水线上,说明蒸发作用使土壤水δ18O发生不平衡分馏,导致δ18O大量富集,这与Wenninger et al[26]研究结果一致.根据Iso Source模型计算各潜在水源对植物水分的贡献比例[27],判别生境上发育植被不同对各层位土壤水以及表层岩溶水的利用差异,从而确定植被对土壤水分再分配的影响,进一步分析研究区植被水分利用对土壤水δ18O的影响.
3.2 喀斯特地区各小生境土壤水运移差异
岩溶区降水主要损失方式为深层渗漏和壤中流,其次为表层岩溶带侧渗和地表径流,这种多界面水文过程共同驱动地表、地下双层土壤侵蚀,养分流失.岩溶区本身成土速率慢,导致石漠化问题突出[24].降水对土壤水补给分为分散面状入渗和集中灌入式补给,后者在岩溶区十分普遍,上坡位岩石裸露,缺少土壤缓冲,会存在灌入式补给[28].岩溶季节性干旱使得表层土壤水供给不稳定,石沟灌丛植被必须通过扩展根幅获取充分的生态空间,侧根发育使石沟土壤容重小,孔隙度大,水分状况良好,这与郭小娇等[12]的研究结果一致.土壤水δ18O季节变化大,优先流运移方式在全年均有体现,旱季降水少,以优先流形式入渗可实现对土壤水的迅速补给,有助于灌丛植物对水源和养分的需求,促进其生长.根据地下水δ18O范围可知地下水也参与了土壤水的补给,雨季小叶羊蹄甲对中层土壤水和表层岩溶水的利用较多,根系发挥水力提升作用,吸收表层岩溶水释放在浅土层使土壤水δ18O值降低.
土面植被优势种为清香木,各月土壤水δ18O均呈波动变化,活塞流推移特征明显.旱季土壤水δ18O总体变化趋势为表层富集,中层波动较大至深层逐渐收缩,可见表层土壤水受蒸发影响强烈,中上层是植被主要的用水层位,深层受地下水影响增大,土壤水δ18O稳定且贫化.有机质、黏粒是土壤团聚体形成的物质基础,有机质含量较高,黏粒较少时,有机质在团聚体形成中起主导作用[29].各生境土壤有机质含量遵循随土层加深迅速衰减的规律,表层含量较高,土壤团聚体结构稳定,土壤透水、蓄水性能良好,抗旱抗冲刷能力强,土壤水文结构功能较好.土面植被优势种清香木表层根系密度高达97.29%,土壤有机质和含水率最高,将植物根系分布与土壤水分峰值对比发现,吸水根系分布深度位于土壤水分峰值范围内,即吸水根系分布区域土壤水分含量最高,旱季利用浅层土壤水较多,雨季利用表层岩溶水较多,表明其土壤水文涵养功能较好.
有机质含量低、黏粒较多时主要靠内聚力形成团聚体,但该类团聚体结构性差,遇雨易崩解,分散的黏粒随壤中流入渗堵塞土壤孔隙,入渗率降低,持水能力弱,易干旱[29].黏粒较重会阻碍植物对土壤水提取[17],各生境土壤黏性均较强,但作为石漠化恢复树种的云南松其主根深至1 m以上,可利用深层土壤水和表层岩溶水,细根主要分布在上层,可有效利用表层土壤水,这种二态根系可从多个层位获得水源,帮助其顺利度过干旱期,将云南松与原生林混交,干旱季节原生林通过水力提升将表层岩溶泉和深层土壤水释放在土壤上层,可以更好的适应喀斯特地区的季节性干旱,提高水源涵养能力[30].
土坡因其地表枯落物多,植被盖度大,下层土壤物理性黏粒含量高,可有效形成土壤团聚体,减弱蒸发作用,云南松对表层岩溶水的水力提升也使土壤水δ18O季节变化减小且贫化,使含水率呈现随深度增加逐渐升高的趋势.旱季各月土壤水δ18O曲线呈波动变化,上层土壤水δ18O变化较小,深层差异较大,可见在旱季云南松不仅可利用表层土壤水,也可通过深根系利用地下水进而影响土壤水的运移.雨季土壤水δ18O呈逐渐收缩趋势,云南松主要利用地下水,使深层土壤水δ18O稳定贫化,有效调节了土壤水分.
4 结论
(1)喀斯特峰林湖盆区土壤水运移不仅受小生境类型差异和土壤水文特征影响,植物群落的水分利用对土壤水再分配也会产生影响.随着各生境上植被群落的恢复,成熟的清香木和云南松林相比于灌丛更趋向于利用表层岩溶水等稳定的水源,土壤水异质性逐渐降低.
(2)石沟小生境雨季表层土壤水δ18O变异程度大,旱季表层和深层土壤水δ18O变异程度较低,混合作用较强,土壤水活塞流和优先流并存,以优先流补给为主;土面雨季各层土壤水变异程度高,基质流混合作用较弱,旱季变异程度较低,深层出现最低值,运移模式多为活塞流;土坡雨季土壤水δ18O表层变异系数大于深层,旱季深层土壤水δ18O变异程度为各生境最大,其它各层均较小,基质流混合作用强,以活塞流运移为主,上层存在优先流.
(3)喀斯特地区植被恢复应选择合适的小生境和植被恢复模式.土壤水分主要来源于大气降水,在全球气候变化背景下,降水丰缺会对喀斯特石漠化地区土壤水分动态变化和群落稳定性产生影响,采取的上坡位灌丛,中坡位次生林,下坡位云南松林的植被恢复模式使土壤水文涵养功能有较好恢复,石漠化发生率降低,表明该植被恢复模式对喀斯特石漠化的治理起到了积极作用.
