麦类作物含R基因组3个物种核型及进化关系分析
2020-07-09任学军李明昊杨梦涵郭振清林小虎
姚 锐, 任学军, 李明昊, 杨梦涵, 郭振清, 高 慧, 林小虎
(河北科技师范学院农学与生物科技学院, 河北 秦皇岛 066004)
麦类作物作为禾本科作物中基因组多倍化较为复杂的一个类群,它们在多倍性进化过程中极易发生染色体重排,广泛的染色体重排增加了基因组的不稳定性,导致了新核型和子代新物种的形成[1]。随着“中国春”小麦基因组测序完成及其近缘物种(粗山羊草、二穗短柄草等)基因组草图的公布,利用比较基因组学方法研究小麦基因组序列组装、染色体结构变异受到许多学者的关注。马建等[2]鉴定了“中国春”小麦4 A、5 A 和7 B易位染色体上参与易位和倒位片段的断点侧翼基因序列;Wang等[3]揭示了小麦、大麦、粗山羊草和短柄草基因组独立的重组关系;孙桑蓉[4]明确了全基因组加倍对于麦类基因家族扩增有着重要作用,并通过生物信息学方法进行了基因组结构解析和多倍化过程的重构。麦类作物基因组草图测序工作的完成为阐明麦类作物基因组的多倍性进化历程起到推动作用,但染色体核型分析在近缘多倍性物种进化关系分析中依然具有重要的理论和现实意义[5-6]。
黑麦属(SecaleL.)是小麦的三级基因源,具有抗白粉病、抗三锈病、抗旱、抗寒等优良性状[7-8]。为了将黑麦中的有益基因转移到小麦中,育种家利用远缘杂交等手段创制了一批小麦-黑麦双二倍体、非整倍体。其中六倍体小黑麦和八倍体小黑麦作为创造小麦-黑麦异附加系、异代换系、异易位系的重要材料,在育种工作中具有重要地位[9]。六倍体小黑麦(AABBRR,2 n=42)由四倍体小麦(T.turgidum, AABB,2 n=28)或普通小麦(TriticumaetivumL. AABBDD,2 n=42)和二倍体黑麦(SecalecerealL. RR,2 n=14)远缘杂交、人工染色体加倍获得,八倍体小黑麦(AABBDDRR,2 n=56)是由普通小麦(TriticumaetivumL. AABBDD,2 n=42)和二倍体黑麦远缘杂交、人工染色体加倍获得。关于二倍体黑麦和六倍体小黑麦的核型已有相关报道,普遍认为二倍体黑麦有14条染色体,但不同品种及生境的二倍体黑麦染色体核型类型不尽相同[10-12];六倍体小黑麦有42条染色体,武晶研究得出的核型分析结果为2 n=42=30 m(2 SAT)+2 M+8 sm+2 st(2 SAT)[13],然而尚未见八倍体小黑麦的核型报道。本研究首次报道了八倍体小黑麦的核型,丰富了小黑麦的细胞遗传学研究,并且将八倍体小黑麦与二倍体黑麦、六倍体小黑麦的核型进行了进化趋势分析,旨在为3个物种的染色体组学研究和相关的麦类作物育种提供细胞学基础。
1 材料与方法
1.1 材 料
供试材料二倍体黑麦(SecalecerealL.)为荆州黑麦;六倍体小黑麦(×TriticalehexaploideL.)由山东农业大学自墨西哥国际玉米小麦改良中心引进;八倍体小黑麦(×TriticaleoctoploidL.)由南京农业大学亓增军教授以荆州黑麦和普通小麦辉县红创制,山东农业大学引进。上述材料均由山东农业大学农学院国家小麦改良中心泰安分中心王洪刚教授课题组提供。
1.2 方 法
1.2.1实验方法
将3种供试材料种子置于盛有蒸馏水的培养皿中室温浸种至露白,移置于湿润的滤纸上室温培养,待根长至1~2 cm时,剪取根尖放入EP管中,冰水浴36 h。用卡诺固定液(乙醇∶冰醋酸体积比3∶1)固定2 d。蒸馏水漂洗后于1 mol·L-1HCl溶液中60 ℃解离8 min,蒸馏水漂洗10 min,改良卡宝品红染液染色2 min,压片并在OLYMPUS BX 53型显微镜下镜检,统计各细胞染色体数目,选取染色体分散良好的细胞拍照。采用Photoshop CS 6软件分析图像[14],并用Excel软件处理数据。
1.2.2染色体核型分析方法及进化指数赋值方法
每个物种选取5个染色体处于有丝分裂中期且分散良好的细胞,参照李懋学等[15]的染色体核型方法计算核型数值。核型类型参照Stebbins[16]的标准。核型不对称系数参照Arano[17]的方法计算。染色体相对长度比、平均臂比、不对称系数、臂比大于1.7的染色体比例采用分支系统学的常规编序、赋值方法进行赋值[18],最终求得可以反映每个物种进化程度的进化指数。具体赋值方法如下:
1) 染色体长度比越大,核型越倾向于不对称。3个物种的染色体长度比可粗略分为4个进化阶段,即1.00以下(赋值为0)→1.00~1.50(赋值为1)→1.51~2.00(赋值为2)→2.01及以上(赋值为3)。
2) 平均臂比值越大,核型越显不对称。3个物种的染色体平均臂比值分为4个阶段,即1.70以下(赋值为0)→1.70~1.90(赋值为1)→1.91~2.10(赋值为2)→2.11及以上(赋值为3)。
3) 臂比大于1.7的染色体比例越大,核型越显不对称。3个物种的染色体臂比大于1.7的染色体比例分为4个进化阶段,即0.20及以下(赋值为0)→0.21~0.40(赋值为1)→0.41~0.60(赋值为2)→0.61及以上(赋值为3)。
4) 不对称系数偏大,核型也显示不对称。3个物种染色体不对称系数分为4个进化阶段,即60以下(赋值为0)→60~62(赋值为1)→62~64(赋值为2)→64以上(赋值为3)。
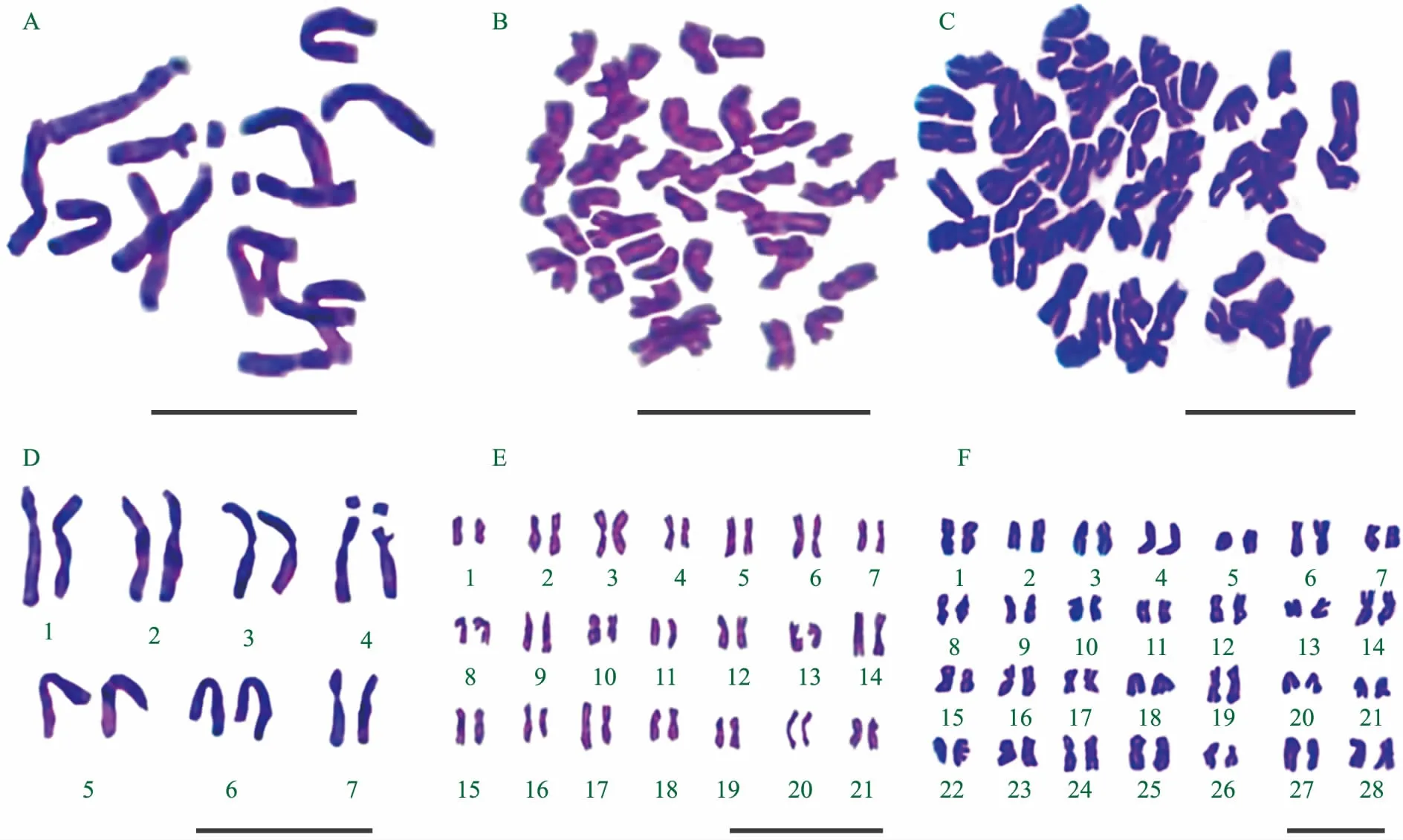
注:A为二倍体黑麦染色体形态;B为六倍体小黑麦染色体形态;C为八倍体小黑麦染色体形态;D为二倍体黑麦染色体核型;E为六倍体小黑麦染色体核型;F为八倍体小黑麦染色体核型。标尺200 μm。
2 结果与分析
本实验共测量分析麦类作物含R基因组3个物种的核型,根尖细胞染色体形态、核型见图1,染色体核型参数及进化指数见表1和表2。
二倍体黑麦染色体相对长度范围为10.36%~16.92%,最长和最短的染色体比值为1.63,臂比大于2∶1的染色体为0对,应属于“1 A”型。核型不对称系数为58.08%,属于较为对称类型。第3、6对染色体为近中部着丝点染色体,其余为中间着丝点染色体,1染色体带有随体,其核型公式为2 n=2 x=14=10 m+4 sm+(2 SAT)(表1)。
六倍体小黑麦染色体相对长度范围为3.50%~6.02%,最长和最短的染色体比值为1.72,臂比大于2∶1的染色体为2对,应属于“2 A”型。核型不对称系数为57.91%,属于较为对称类型。第12对为正中部着丝点染色体,第3、11、15对染色体为近中部着丝点染色体,其余为中部着丝点染色体,其核型公式为2 n=6 x=42=2 M+34 m+6 sm(表1)。
八倍体小黑麦染色体相对长度范围为2.45%~4.62%,最长和最短的染色体比值为1.89,臂比大于2∶1的染色体为1对,应属于“2 A”型。核型不对称系数为56.44%,属于较为对称类型。第15、23、25对染色体为正中部着丝点染色体,第14、24、26对染色体为近中部着丝点染色体,其余为中部着丝点染色体,其核型公式为2 n=8 x=56=6 M+44 m+6 sm(表1)。
依据Stebbins提出的核型进化理论,核型进化是由对称向不对称方向发展的[13]。核型参数可以反映每个物种发育的进化程度,从而可以反映各自的进化水平。核型为1 A的二倍体黑麦进化程度较为原始,核型为2 A的六倍体小黑麦和八倍体小黑麦进化程度相对较高。由表2可知,二倍体黑麦、六倍体小黑麦、八倍体小黑麦进化指数分别为3、4、4,表明六倍体小黑麦和八倍体小黑麦进化程度相同且高于二倍体黑麦的进化水平。
3 讨 论
本研究的2种小黑麦是由四倍体小麦(AABB,2 n=28)或普通小麦(AABBDD,2 n=42)和二倍体黑麦(RR,2 n=14)杂交培育获得的双二倍体,带有亲本A、B、(D)、R基因组。研究表明,小麦族B基因组中,1 B、6 B染色体带有随体[19];二倍体黑麦1 R染色体带有随体[10],但也有不同观点,代明等[12]、Gill[20]的研究指出,有些黑麦染色体不带有随体,不同居群、生境、品种的黑麦染色体核型存在差别。小黑麦是人工合成的作物,含有亲本2个不同物种的染色体,染色体遗传机制较为复杂,理论上六倍体小黑麦和八倍体小黑麦都应该有6条染色体带有随体。然而,2种小黑麦根尖细胞观察结果显示,没有带6条随体染色体的细胞出现。在部分六倍体小黑麦根尖细胞中,观察到带1条随体染色体的情况,说明本试验所采用的六倍体小黑麦材料,至少应该有2条染色体带有随体;在部分八倍体小黑麦根尖细胞中,观察到带有1条随体、2条随体、3条随体染色体的情况,说明本实验所采用的八倍体小黑麦材料可能有4条或6条染色体带有随体。在武晶[13]的研究中,由四倍体硬粒小麦和二倍体黑麦培育的六倍体小黑麦Occurance 54-22核型公式为2 n=42=30 m(2 SAT)+2 M+8 sm+2 st(2 SAT),含有普通小麦A、B基因组和黑麦R基因组,其中4条染色体带有随体,与理论上的6条带随体染色体数目不符。在本研究中,2种小黑麦材料体细胞中同样未见与理论相符的带随体的染色体数目。
表1 麦类作物含R基因组3个物种染色体参数
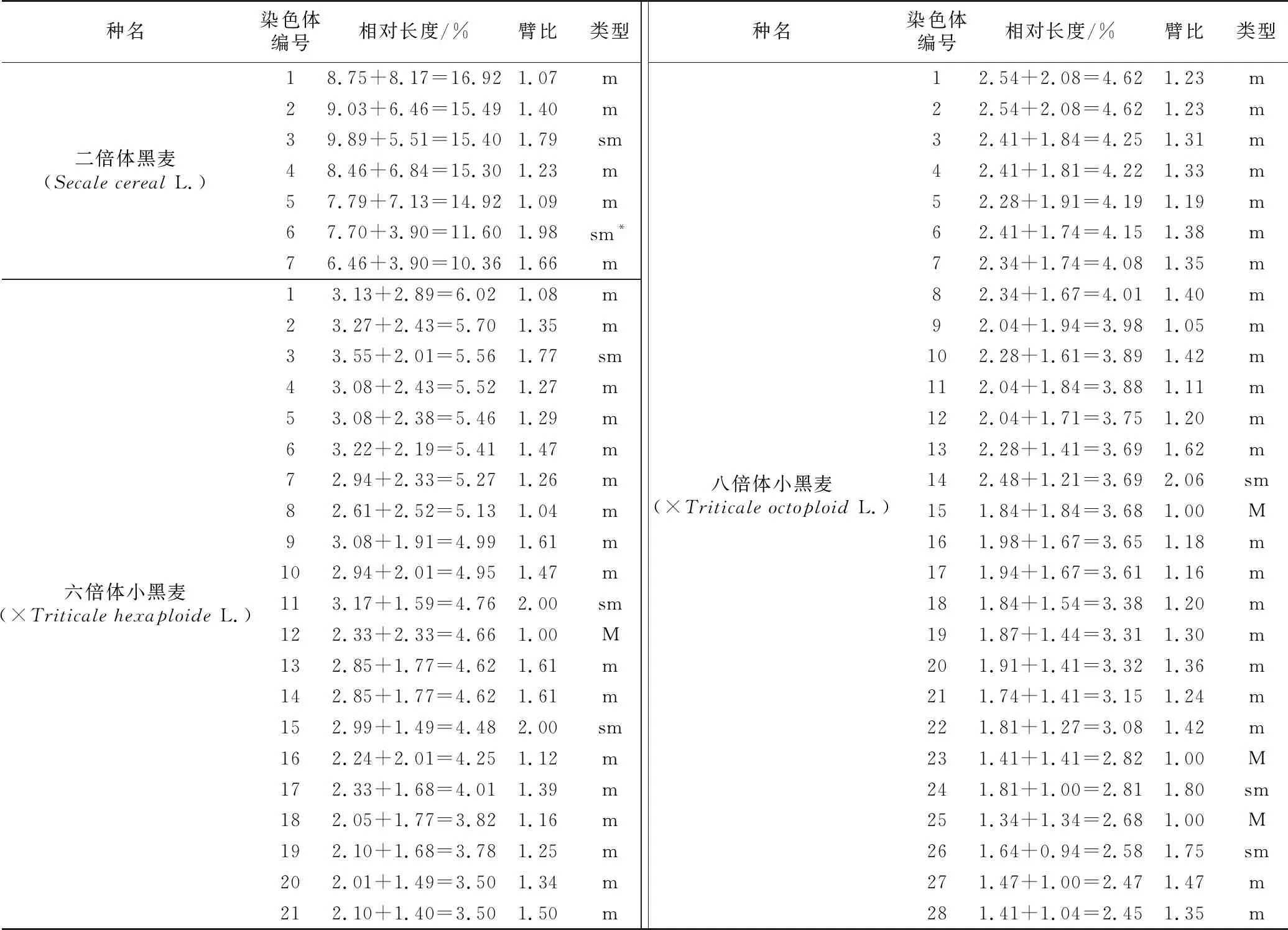
种名染色体编号相对长度/%臂比类型二倍体黑麦(Secale cereal L.)18.75+8.17=16.921.07m29.03+6.46=15.491.40m39.89+5.51=15.401.79sm48.46+6.84=15.301.23m57.79+7.13=14.921.09m67.70+3.90=11.601.98sm*76.46+3.90=10.361.66m六倍体小黑麦(×Triticale hexaploide L.)13.13+2.89=6.021.08m23.27+2.43=5.701.35m33.55+2.01=5.561.77sm43.08+2.43=5.521.27m53.08+2.38=5.461.29m63.22+2.19=5.411.47m72.94+2.33=5.271.26m82.61+2.52=5.131.04m93.08+1.91=4.991.61m102.94+2.01=4.951.47m113.17+1.59=4.762.00sm122.33+2.33=4.661.00M132.85+1.77=4.621.61m142.85+1.77=4.621.61m152.99+1.49=4.482.00sm162.24+2.01=4.251.12m172.33+1.68=4.011.39m182.05+1.77=3.821.16m192.10+1.68=3.781.25m202.01+1.49=3.501.34m212.10+1.40=3.501.50m种名染色体编号相对长度/%臂比类型八倍体小黑麦(×Triticale octoploid L.)12.54+2.08=4.621.23m22.54+2.08=4.621.23m32.41+1.84=4.251.31m42.41+1.81=4.221.33m52.28+1.91=4.191.19m62.41+1.74=4.151.38m72.34+1.74=4.081.35m82.34+1.67=4.011.40m92.04+1.94=3.981.05m102.28+1.61=3.891.42m112.04+1.84=3.881.11m122.04+1.71=3.751.20m132.28+1.41=3.691.62m142.48+1.21=3.692.06sm151.84+1.84=3.681.00M161.98+1.67=3.651.18m171.94+1.67=3.611.16m181.84+1.54=3.381.20m191.87+1.44=3.311.30m201.91+1.41=3.321.36m211.74+1.41=3.151.24m221.81+1.27=3.081.42m231.41+1.41=2.821.00M241.81+1.00=2.811.80sm251.34+1.34=2.681.00M261.64+0.94=2.581.75sm271.47+1.00=2.471.47m281.41+1.04=2.451.35m
注:M、m、sm、*分别表示正中部着丝点染色体、中部着丝点染色体、近中部着丝点染色体、随体,随体长度未计算在内。
表2 麦类作物含R基因组3个物种染色体重要性状及编码
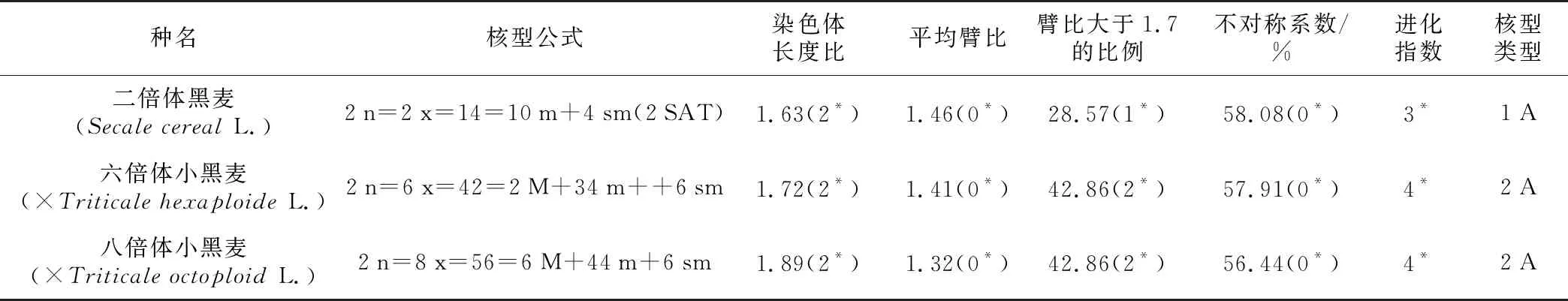
种名核型公式染色体长度比平均臂比臂比大于1.7的比例不对称系数/%进化指数核型类型二倍体黑麦(Secale cereal L.)2n=2x=14=10m+4sm(2SAT)1.63(2*)1.46(0*)28.57(1*)58.08(0*)3*1A六倍体小黑麦(×Triticale hexaploide L.)2n=6x=42=2M+34m++6sm1.72(2*)1.41(0*)42.86(2*)57.91(0*)4*2A八倍体小黑麦(×Triticale octoploid L.)2n=8x=56=6M+44m+6sm1.89(2*)1.32(0*)42.86(2*)56.44(0*)4*2A
注:“*”为性状状态编码数值。
目前,关于植物细胞随体染色体的研究不够深入,随体的遗传机制尚不明确。在禾本科植物中,普遍认为水稻有2对带随体染色体,然而不同品种、不同生境的水稻有1对、2对、3对随体染色体的报道[21,22],尚未发现相关的研究成果对这一现象进行解释,可能是因为染色体随体伸长或者随体染色体的NOR活性不同引起的淡化导致的假象。另外,依据穆尔(莫慧英译)提出的理论,也可能是因为核仁合并或者亲本中一个种的核仁中心对另一个种呈显性导致随体消失[23]。此外,Li等[24]研究发现,二倍体小麦和黑麦在4 L上有相同的染色体重排断点,5 L上的染色体重排断点位置大体相近,表明小黑麦在进化过程中极有可能在4 L5 L处发生相应的异位,这些变化都可能引起同源共线性基因片段的变化或基因组的重排,从而发生染色体片段的倒位、重复、插入和缺失等现象,进而影响染色体间的重组,最终导致麦类作物多倍性进化历程中核型的变化。
在本研究中,二倍体黑麦核型公式为2 n=2 x=14=10 m+4 sm(2 SAT),核型类型为1 A,进化指数为3;六倍体小黑麦核型公式为2 n=6 x=42=2 M+34 m+6 sm,核型类型为2 A,进化指数为4;八倍体小黑麦核型公式为2 n=8 x=56=6 M+44 m+6 sm,核型类型为2 A,进化指数为4,六倍体小黑麦和八倍体小黑麦核型进化程度高于二倍体黑麦。由于小黑麦具有亲本2个不同物种的全套基因,可以利用2个基因库的基因来抵挡环境的变化,在进化层面上,小黑麦应该进化程度更高,在农艺生产中,小黑麦确实兼具黑麦和小麦的优良性状[25],核型分析得出的结论也验证了这一点。
