光质对珙桐幼苗形态建成及光合特性的影响
2020-07-08吴艳,荣熔,陈放,徐莺
吴 艳, 荣 熔, 陈 放, 徐 莺
(四川大学生命科学学院生物资源与生态环境教育部重点实验室, 成都 610065)
1 引 言
在植物的生命周期中,光是对其影响最为显著的环境因子.光照强度、光质以及光周期都会对植物的生长发育、光合作用以及生理生化特性等方面均有较大的调控作用[1-3].
珙桐(Davidiainvolucrata)是我国特有的单型属珍稀树种,有植物界的“活化石”之称,被列为国家一级保护植物.珙桐是一种著名的观赏植物,因其有一对白色的大苞片,形如白鸽的翅膀,所以珙桐又名鸽子树[4-5];此外,珙桐还具有较高的经济价值,其种子和果皮可以用于榨取食用油或工业油、果核可提取营养蛋白质、果内皮可制香精以及树皮果皮可制成活性炭等[6].作为濒危珍稀物种,一方面珙桐对生长环境的要求较为严格[7],;另一方面,珙桐的生长习性尤为特殊:其幼苗生长缓慢,喜阴湿的生长环境,适宜生长在林荫下,成苗趋于喜光,可知光照对珙桐植株的生长有关键影响.早前的研究对珙桐生长产生影响的因子主要有温度[8]、光强[9]等,但有关于光质影响珙桐生长的研究却未见报道.本试验探讨了不同弱光光质对珙桐幼苗生长形态及生理特性的影响,以期为其种质资源的保护、栽培引种技术及园林应用方面提供相关的理论依据.
2 材料与方法
2.1 材 料
本研究的供试植物材料为野生型光叶珙桐,种子采摘于四川省江油市吴家后山.
2.2 方 法
2.2.1 培养条件 本试验选取颗粒饱满的珙桐种子于人工气候培养箱中进行萌发和培养,条件设置为温度 24 ℃,光照强度3 000~4 000 lx,光照周期14 h光照/10 h黑暗,相对空气湿度为80%.
2.2.2 光质处理 本试验一共设置了4种不同LED光质处理,分别为白色(W)、蓝色(B)、红色(R)、绿色(G),光照强度保持一致(3 000 lx),光周期14 h光照/10 h黑暗.每个处理重复3次,每次重复3~5棵苗,培养周期为35 d.
2.2.3 形态指标测定 选取相同位置、方向一致的叶片测定相关的形态指标.以叶片长度大于5 cm算作真叶的有效准则,统计幼苗的真叶数.测定株高、上胚轴长度(子叶与第一片真叶之间的距离)、茎粗(子叶以下1 cm处的部位)、叶长、叶宽、叶柄长度,利用扫描仪(EPSON scan)对幼苗叶片进行扫描,之后再采用Photoshop CS4软件测定叶片面积.
2.2.4 光合色素含量测定 叶绿素含量采用95%乙醇提取法[10]进行测定,利用打孔器将叶片打成直径1 cm的小圆片,称取0.1~0.2 g左右的叶片材料,将材料快速放入装有事先配置好的95%酒精的塑料试管中,暗处理72 h之后至叶片完全不呈绿色,利用酶标仪(Molecular Devices,USA)分别测定波长665 nm、649 nm、440 nm、652 nm下的吸光值,计算得到叶绿素a (Ca)、叶绿素b (Cb)、叶绿素总含量(CT)、类胡萝卜素(Cx.c),
计算公式如下:Ca=(12.7×OD665-2.59×OD649)×L/(1000×M);Cb=(22.9×OD649-4.67×OD665) ×L/(1000×M);CT=Ca+Cb;Cx.c= (1000×OD470-2.05×Ca-114.8×Cb) /245×L/(1000×M).
公式中L:混合液体积(mL),M:样品质量(g).
2.2.5 叶绿素荧光参数测定 将待测的珙桐幼苗经过20 min暗处理之后,MINI-IMAGING-PAM荧光成像仪(德国WALZ 公司)测定叶绿素荧光成像参数PSⅡ最大光量子产量(Fv/Fm)、PSⅡ实际光量子产量Y(Ⅱ)、光化学淬灭系数qP、非光化学淬灭系数qN、PSⅡ调节性能量耗散的量子产量Y(NPQ)、非调节性能量耗散的量子产量Y(NO)及快速光曲线,并通过快速光曲线计算得到初始斜率α、最大潜在电子传递速率rETRm、半饱和光强Ik.
2.2.7 统计学处理 试验数据用EXCEL 2010软件进行整理,利用DPS7.05软件进行显著性分析,Duncan新复极差法进行多重比较.
3 结果与分析
3.1 不同光质对珙桐幼苗形态建成的影响
本研究首先测定了不同光质下的珙桐幼苗及其叶片的形态学参数(表1,图1).在植株形态方面,白光下的珙桐幼苗株高最高,其次是蓝光和绿光,红光处理下株高最小,但是幼苗的上胚轴和茎的直径在蓝光处理下达到最大值.在叶片数量和形态方面,蓝光与白光处理下的幼苗真叶数均显著高于绿光和红光,白光处理下叶片的长度、宽度及叶面积均显著高于其他处理,且叶片最为舒展宽阔,而蓝、红、绿三种处理下的叶片形态依次变得越来越狭长,但叶片长宽比却是在绿光和红光处理下达到最大且显著高于另外两组处理.绿光下幼苗的叶柄长度显著高于其他三组处理,其次是红光,白光和蓝光下叶柄最短.这些结果表明,在不同光质环境中生长时,珙桐幼苗植株的形态建成会对环境条件做出不同的响应.
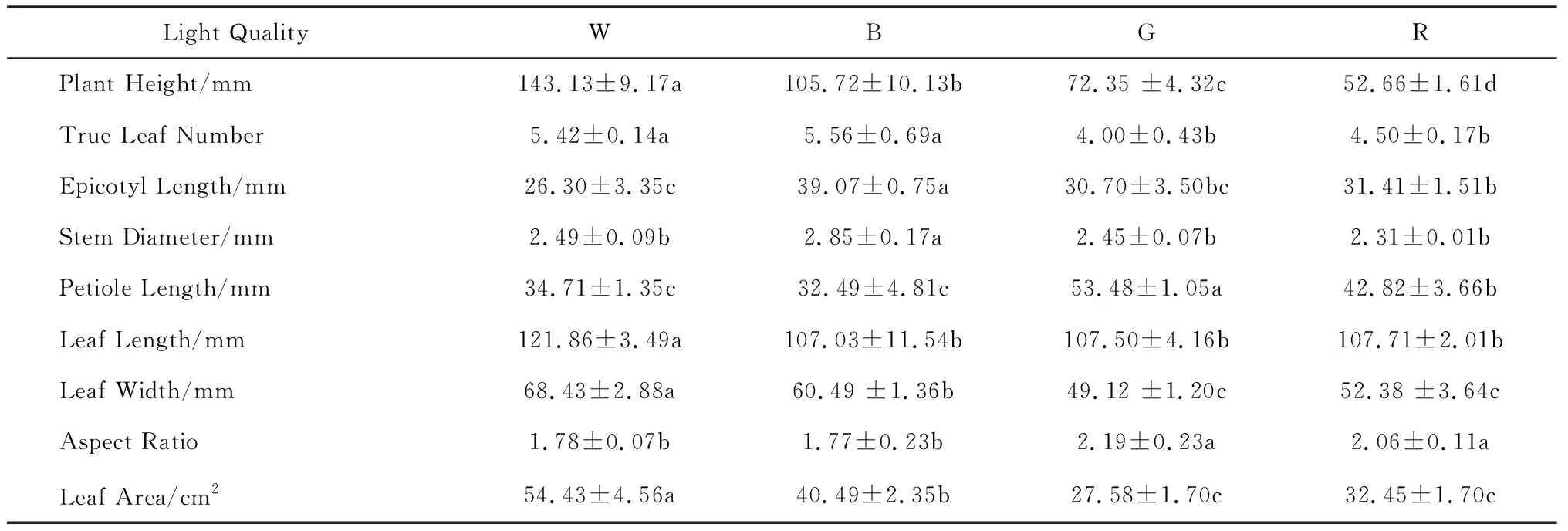
表1 不同光质处理后珙桐幼苗的形态建成指标
(a) 经不同光照处理35 d后的珙桐幼苗; (b) 经不同光照处理35 d后的珙桐幼苗叶片. (W, 白光; B, 蓝光; R, 红光; G, 绿光.)
Fig.1 Plants and leaves morphology of seedlings treated with different light quality
(a) Plants morphology; (b) leaves morphology.(W, white light; B, blue light; R, red light; G, green light.)
3.2 不同LED光质对珙桐幼苗光合色素含量的影响
不同光质条件下,珙桐幼苗叶片的叶绿素总含量、类胡萝卜素含量以及叶绿素a、叶绿素b的含量均呈红光>绿光>白光>蓝光依次递减的趋势,且红光和绿光处理的幼苗色素含量显著高于其他两组处理.而叶绿素a与叶绿素b的比值却与叶绿素a和叶绿素b的含量恰恰相反,呈现出蓝光>白光>绿光>红光依次递减的趋势.这表明红光和绿光更有利于幼苗光合色素的累积和光合作用的进行(图2).
3.3 不同LED光质对珙桐幼苗叶绿素荧光特性的影响
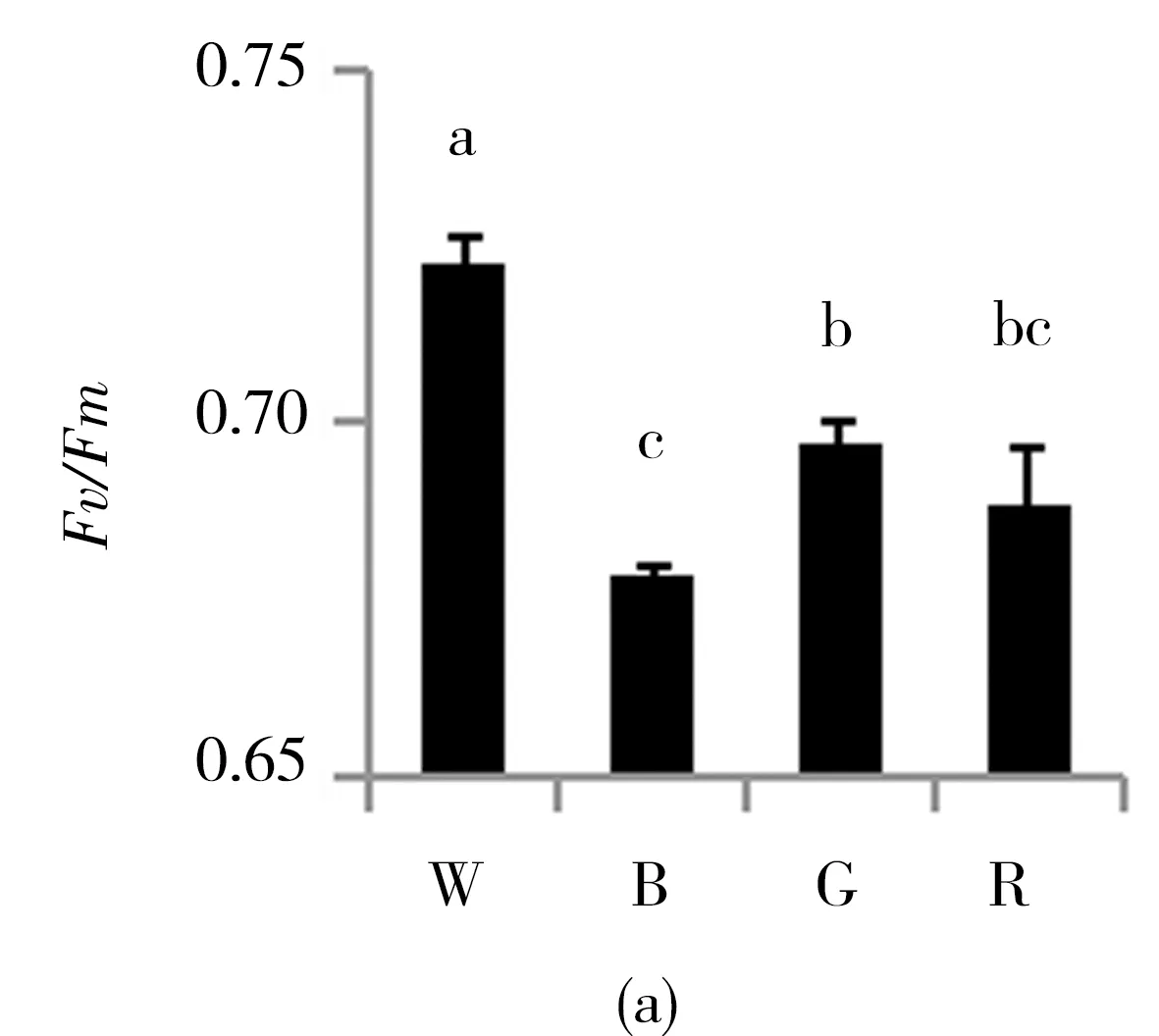
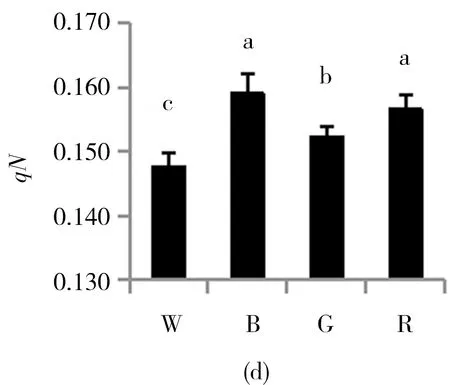
由图3可知,白光处理的珙桐幼苗,其PSⅡ最大量子产量Fv/Fm和PSⅡ实际量子产量Y(Ⅱ)都达到最大值,并且显著高于其他光质处理,而经蓝光处理的幼苗这两项参数值最低.四种不同光质处理后幼苗的光化学淬灭系数qP和非光化学淬灭系数qN呈现出完全相反的两种结果,光化学淬灭系数qP在白光和绿光处理的幼苗中较高,而非光化学淬灭系数qN却在这两种光质处理下较低,反而在蓝光和红光处理的幼苗中较高.不同光质处理后,PSⅡ处调节性能量耗散的量子产量Y(NPQ)和PSⅡ处非调节性能量耗散的量子产量Y(NO)的结果呈一定的正相关性,Y(NPQ)和Y(NO)在蓝光和红光处理下均显著高于白光和绿光处理.
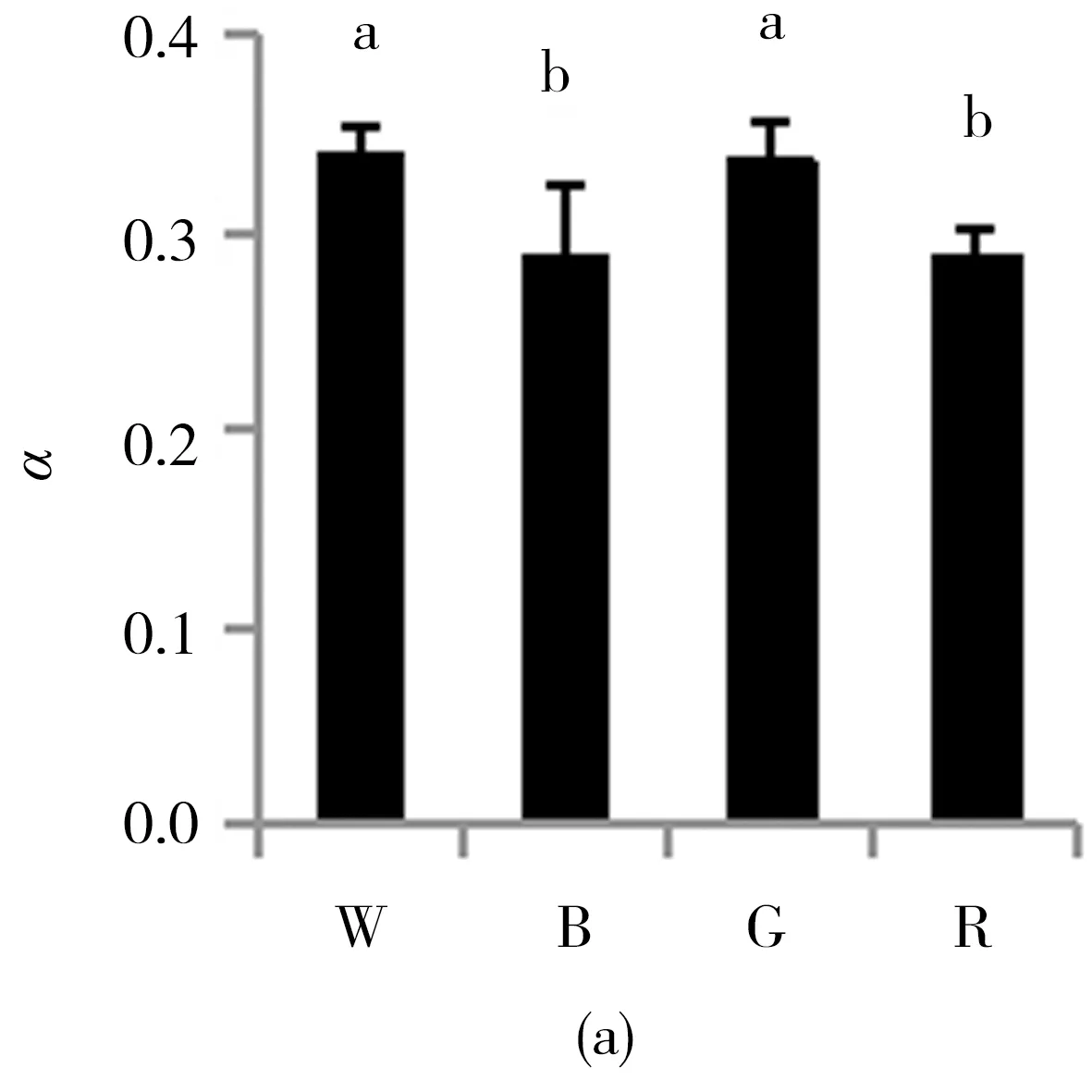
此外,快速光曲线初始斜率α在白光与绿光处理下显著高于蓝光和红光处理,白光和绿光处理间无显著差异,蓝光和红光处理间同样无差异(图4a);半饱和光强Ik在蓝光处理下达到最大值,且显著高于其他处理;其次是绿光和白光处理,两者间不存在差异;红光处理下Ik最低,显著低于其他处理(图4b);最大潜在电子传递速率rETRm白光和绿光处理下较高,且显著高于其他处理,其次是蓝光处理,而红光处理则最低(图4c).
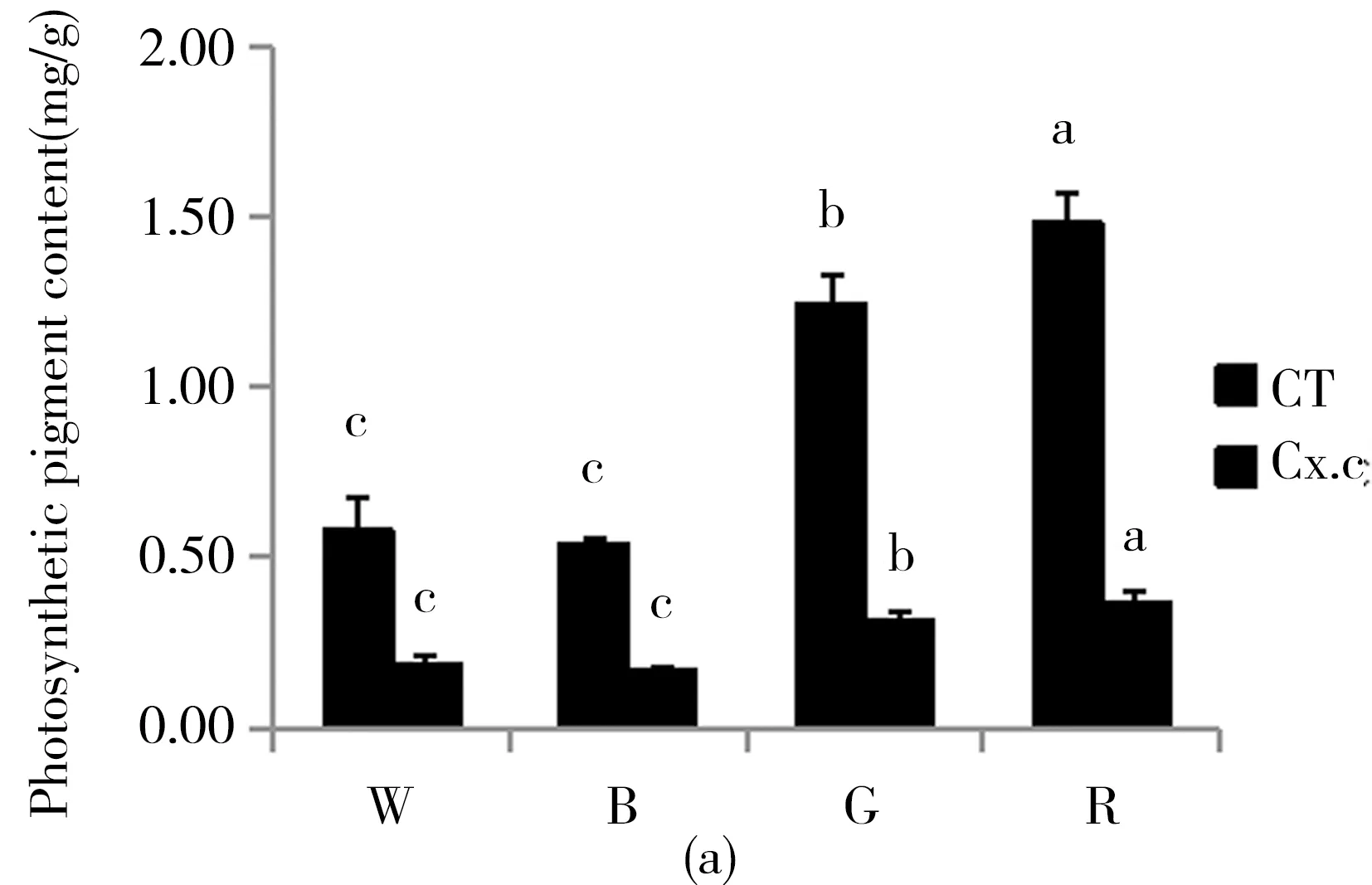
(a) The content of total chlorophyll and carotenoids of treated seedings; (b) the content of chlorophyll a, chlorophyll b and the ratio of chlorophyll a/b.(CT,total chlorophyll;Cx.c,carotenoid;Ca,chlorophylla;Cb,chlorophyll b.)
(a) PSⅡ最大量子产量(Fv/Fm); (b) PSⅡ实际量子产量Y(Ⅱ); (c) 光化学淬灭系数qP; (d) 非光化学淬灭系数qN; (e) PSⅡ处调节性能量耗散的量子产量Y(NPQ); (f) PSⅡ处非调节性能量耗散的量子产量Y(NO).
Fig.3 Chlorophyll fluorescence parameters of seedling leaves treated with different light quality
(a)Fv/Fm(optimal quantum yield of PSⅡ); (b)Y(Ⅱ)(Actual photochemical efficiency of PSⅡ); (c)qP(Photochemical quenching coefficient); (d)qN(Non-photochemical quenching coefficient); (e)Y(NPQ)(quantum yield of energy dissipation by PS Ⅱ regulatory); (f)Y(NO)(Quantum yield of non-regulatory energy dissipation).
(a) The initial slopeαof the rapid light curve; (b) the half-saturated light intensityIk; (c) the maximum potential electron transfer raterETRm.
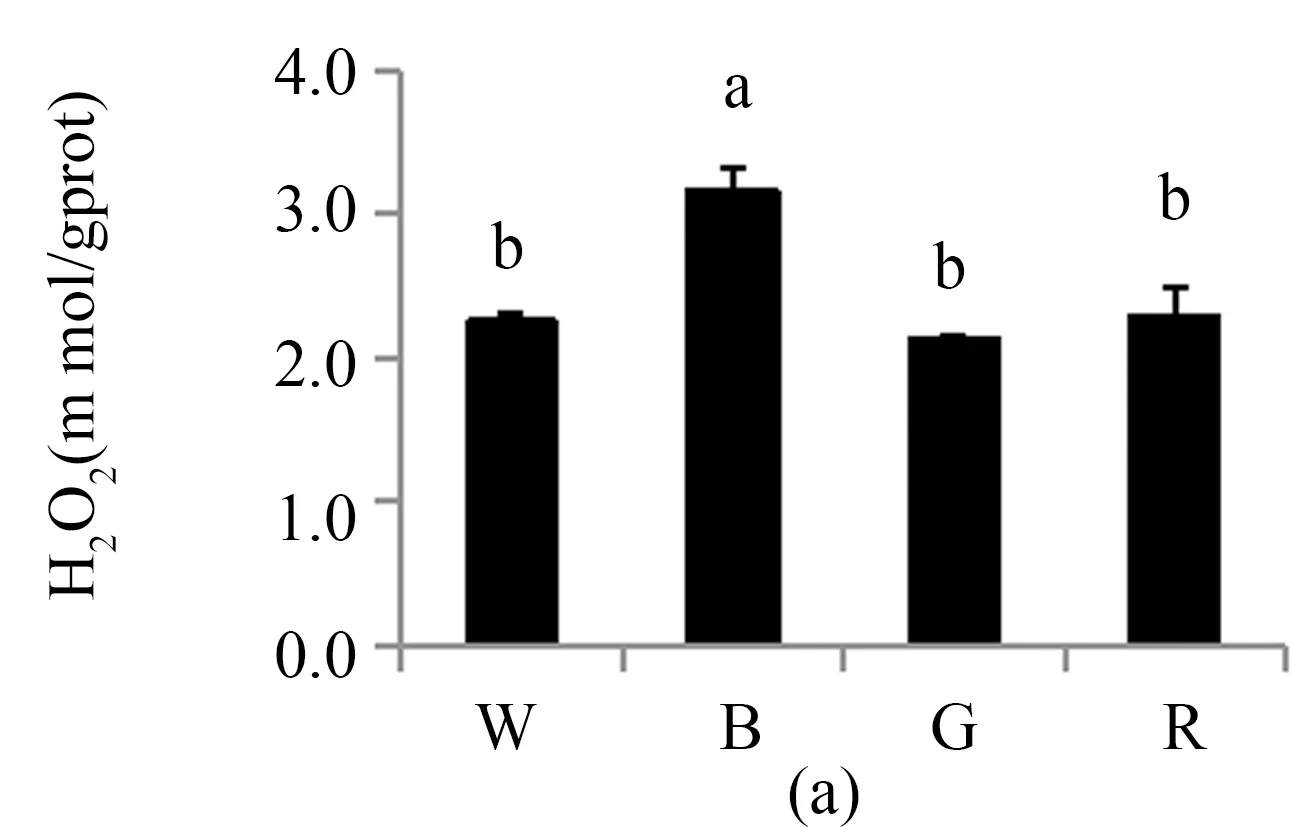
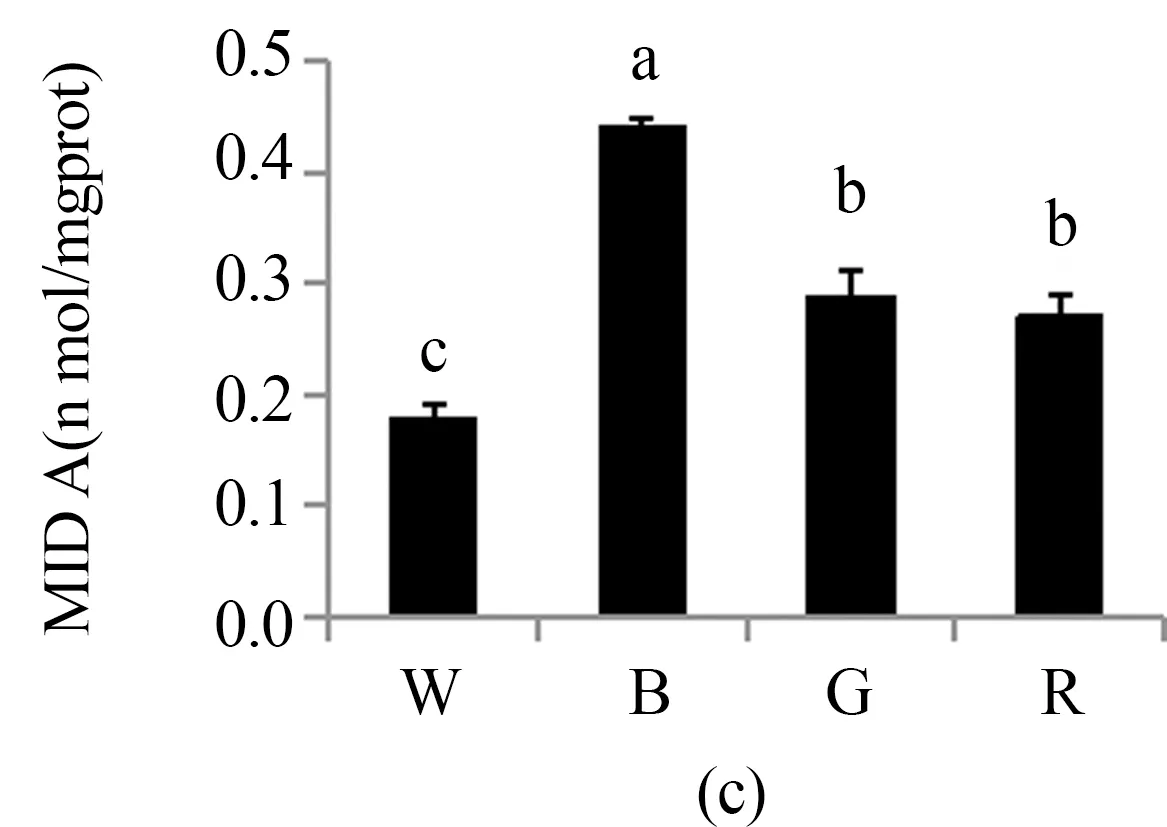
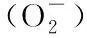
Fig.5 The content of parameters related to reactive oxygen metabolism in seedlings after different light quality treatment
(a) Hydrogen peroxide (H2O2) content; (b) superoxide anion (O2) content; (c) malondialdehyde (MDA) content; (d) SOD activity.
3.4 不同LED光质对珙桐幼苗活性氧代谢的影响
4 讨 论
光照是影响植物形态建成和生长发育的重要外界环境因素。植物不仅能对光照强度做出响应[11],也能对不同光质或者光周期做出应答。作为国家一级保护植物,珙桐的致危因素包括自身内部因素和外界环境因素[12]. 以往的研究主要针对于光照强度对珙桐幼苗生长的影响[9,13],本研究则探讨了白、蓝、绿、红4种光质对珙桐实生幼苗的光形态建成和光合生理特性的影响,结果发现珙桐幼苗对不同光质的响应存在较大差异.
在避荫条件下,被子植物主要表现出加快节间和叶柄的伸长速度、减少叶片区域和厚度、减少分枝、提前开花等避荫反应[14-15].本研究中红光处理的珙桐幼苗株高值最低,说明红光最有利于珙桐幼苗的光形态建成,这与此前报道的研究结果一致[16]. 蓝光对植物的伸长生长具有抑制作用, 但本实验中蓝光处理的珙桐幼苗株高相对较高,可能是蓝光引起了珙桐幼苗的避荫效应而导致.研究发现,绿光可以诱导拟南芥叶柄伸长的避荫应答[17],在本研究中绿光处理的珙桐幼苗叶柄长度显著长于其他处理,同时该处理下珙桐幼苗的叶长、叶宽及叶面积均达到最小值,且叶长与叶宽的比值最高,狭长的珙桐幼苗叶片同样说明绿光处理诱发了避荫效应.研究指出,适宜珙桐幼苗生长的光照强度为10 000~17 000 lx[18],而本研究所采用的试验光强(3 000 lx)远远达不到该水平.该强度的白光下的珙桐幼苗最高,这可能是由于红光仅占其中的一小部分,绿光和低强度蓝光又具有诱导避荫反应能力共同作用的结果.研究发现的在3 000 lx的各种光质处理下的珙桐幼苗叶片面积较生长在光强为10 000 lx的环境中的幼苗叶片面积低,这与早前报道的植物叶片在弱光环境下叶面积下降一致. 这些结果表明,在不同弱光光质处理下,珙桐幼苗叶片的生长发育受到一定的影响,但其中不同光信号的具体分子调控机制还有待进一步研究.
光合色素是绿色植物光合作用的基础.一般而言,蓝光有利于提高叶绿素a与叶绿素b的比值,而蓝光培养的植株一般具有阳性植物的特性,而红光培养的植株与阴性植物相似[19].本实验中在蓝光处理下珙桐幼苗叶绿素a与叶绿素b的比值达到最大值,这说明珙桐是一种典型的阳性植物. 在红光和绿光处理后,珙桐幼苗叶绿素a与叶绿素b的比值下降,表明珙桐幼苗具有一定的耐荫能力,可以通过提高叶绿素b的相对含量来捕获有限的光能来适应弱光环境.Fv/Fm反映植物的潜在最大光合能力,Y(Ⅱ)反映植物的实际光合能力[20]. 本研究结果显示在白光处理下,珙桐幼苗的Fv/Fm和Y(Ⅱ)均达到最大值,说明在弱光条件下,白光仍能保证珙桐幼苗的光合能力,而其他处理下珙桐幼苗的光合能力都有不同程度的下降,其中蓝光处理下的珙桐幼苗光合能力显著低于红光和绿光处理,这与前期的研究结果一致[21]. 一方面,蓝光有利于提高光合电子传递链PSⅠ受体端的自动氧化酶类活性,促进了Mehler反应从而会生成更多的超氧化物[22],本研究中实验结果与此相符;另一方面,蓝光处理下珙桐幼苗叶片的ROS水平最高,导致其MDA含量也同样显著高于别的处理.此外,蓝光下珙桐幼苗可能具有SOD机制之外的抗氧化自我修复能力,能保证植株新陈代谢正常运行,所以蓝光下珙桐幼苗的形态生长状态仍然较佳.
综上所述,在不同光质条件下,珙桐幼苗的形态特征和生理特征存在着显著差异,即使在同一种光照条件下,各种形态和生理指标的响应效果也不尽相同.具体表现为珙桐幼苗在红光处理下伸长生长最缓慢,光合色素累积最高而光和能力最低,这表明红光对植株的光形态建成有显著影响;白光处理的幼苗株高最高,光合色素积累少,但光和能力最强,此外ROS水平及MDA含量较低,且SOD活性达到最大值,说明白色弱光处理对珙桐幼苗的光合作用及生理代谢影响较小;弱光环境下绿光和蓝光处理的珙桐幼苗都能表现出避荫效应,这对于珙桐幼苗的生长发育有一定的积极作用.
珙桐是珍稀濒危的木本植物,实验材料数量的局限使得实验也受到诸多限制,以后还需要在有限的实验材料的前提下,从形态解剖学和基因表达的角度探讨珙桐的光响应;此外,从分子水平上去对其进行分析研究光质对珙桐影响的具体机制也更具有说服力.
