海拔梯度对山噪鹛鸣声的影响
2020-07-03李奇龙
张 雷,李奇龙,唐 毅
(辽宁大学 生命科学院,辽宁 沈阳 110036)
0 引言
鸣声是鸟类重要通讯手段,在吸引配偶、保卫领域、警戒、躲避捕食行为中发挥着重要作用[1].声学分析是揭示鸟类鸣声特征的重要技术,利用该技术曾发现鸣声复杂程度与鸟类多样性之间存在正相关关系[2,3].随着野外录音设备和分析技术的完善,鸣声的声学分析已成为鸟类行为与进化研究的重要方面[4].
环境因素是鸟类鸣声特征分化的重要驱动力.在低噪音背景下的城市或河流区域中,鸟类往往会提高鸣声频率[5-7],以促进鸣声有效传播,这表明鸟类可通过差异性鸣声提高声音传播效率.纬度、降水等地带性因素对鸣声特征亦有影响.喜马拉雅雀(Himalayanwarbler)的鸣声复杂性随纬度梯度增加而变化[7];降水变异导致北美雀形目鸟类鸣声复杂性变化[8].海拔梯度差异可导致温度、降水、植物物种多样性变化,进而可能影响到鸟类鸣声.鸟类的鸣声特征如何受海拔梯度影响,尚不完全清楚.
山噪鹛(Garrulaxdavidi)隶属于雀形目鹟科,为我国特有种,共分为4个亚种,在我国分布较为广泛,南端可到四川中部、西界为甘肃西北部、东北界为辽宁和内蒙古.山噪鹛是我国噪鹛属中分布最北的物种.山噪鹛分布区域海拔变异较大,具有复杂的鸣唱行为和出色的效鸣能力[9],是研究海拔梯度对鸟类鸣声的理想目标.
本研究以山噪鹛为研究对象,计算声学特征,进而分析海拔梯度对其鸣声的影响.本研究有助于揭示海拔梯度对山噪鹛鸣声的影响规律,加深对鸣禽响应环境变化的理解.
1 材料与方法
1.1 数据来源
本研究数据来源于记录世界范围内鸟类鸣声数据的网站https://www.xeno-canto.org/.检索关键词Garrulaxdavidi得到15条记录.删除一条不含海拔信息的记录,最终保留14条可用记录(表1).

表1 山噪鹛鸣声数据记录
1.2 数据处理与分析
本研究分析鸣声声学特征,计算指标包括声学复杂度指数、声学多样性指数、声音均匀度指数、声学丰富度、生物声学指数、声学熵、振幅指数、标准声景差异指数、频峰数.各声学指标含义见表2.以上指标的计算利用R语言程序包turnR、soundecology和seewave完成[10].
本研究探索海拔梯度与鸣声的关系.以海拔高度为预测变量,以鸣声指标为响应变量,构建回归模型.计算回归方程的校正决定系数、回归参数预测值与标准误差、t统计量及相应P值.以上计算使用R语言完成[10].

表2 生物声学系列指标
2 结果
2.1 鸣声声学特征

图1 声学指标箱式图
声学复杂度指数最大值、最小值为183.7、162.2,分别发生在海拔2 600 m 与300 m处,声学复杂度指数均值(±标准差,下同)为171±6.22.声学多样性指数最大值、最小值为2.15、0.03,出现在海拔2 600 m与3 600 m处,其均值为1.45±0.56.声学均匀度指数最大值、最小值为0.9、0.3,出现在海拔3 600 m与2 600 m处,其均值为0.62±0.18.声学丰富度最大值、最小值为0.768、0.003,发生在海拔 3 400 m与150 m处,其均值为0.23±0.22.生物声学指数最大值、最小值为66.1、12.4,出现在海拔300 m与2 600 m处,其均值为35.1±17.3.声学熵最大值、最小值为0.82、0.30,出现在海拔340 m与3 600 m处,其均值为0.64±0.16.振幅指数最大值、最小值为0.12、0,出现在海拔3 400 m与150 m处,其均值为0.032±0.033.标准声景差异指数最大值、最小值为0.96、-0.11,出现在海拔150 m与2 600 m处,其均值为0.71±0.30.频峰数最大值为3,表明山噪鹛鸣声至多存在3个频峰;海拔高于3 000 m区域,鸣声频峰数均为0,表明无频峰(图1).
2.2 鸣声特征与海拔关系
声学多样性指数、声学均匀度指数、声学熵与海拔梯度分别存在显著的函数关系(表3).声学多样性指数与声学熵随海拔梯度增加先升高后降低,其最大值分别出现在海拔1 718 m和1 646 m处(图2(a)、(c)).声学均匀度指数随海拔梯度增加先降低后升高,最小值产生在海拔1 783 m处(图2(b)).其他指标与海拔梯度之间未发现显著的函数关系(表3).
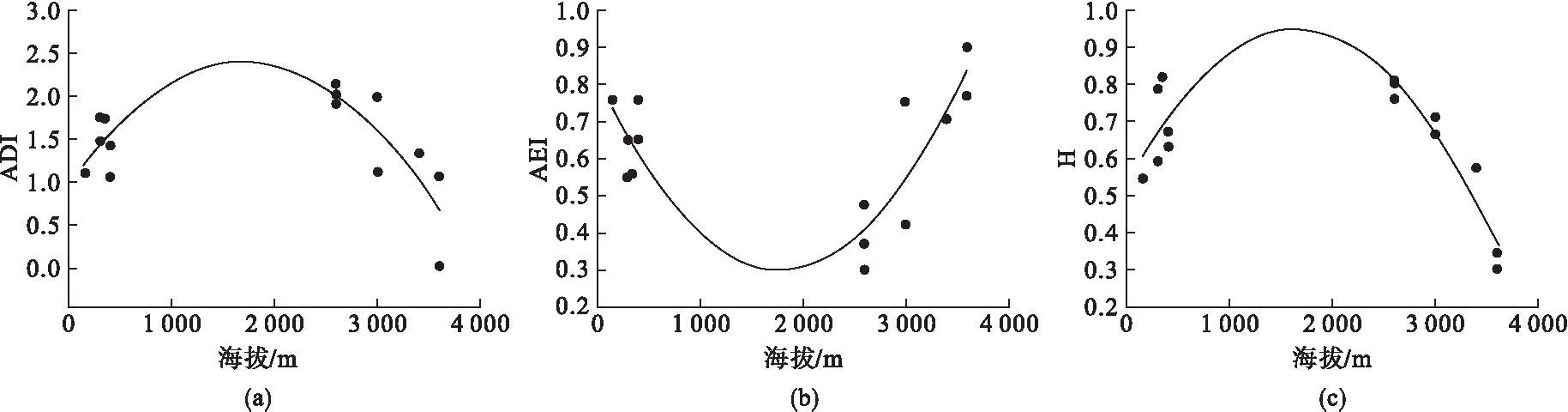
图2 海拔梯度与声学多样性指数(a)、声学均匀度指数(b)、声学熵(c)的关系
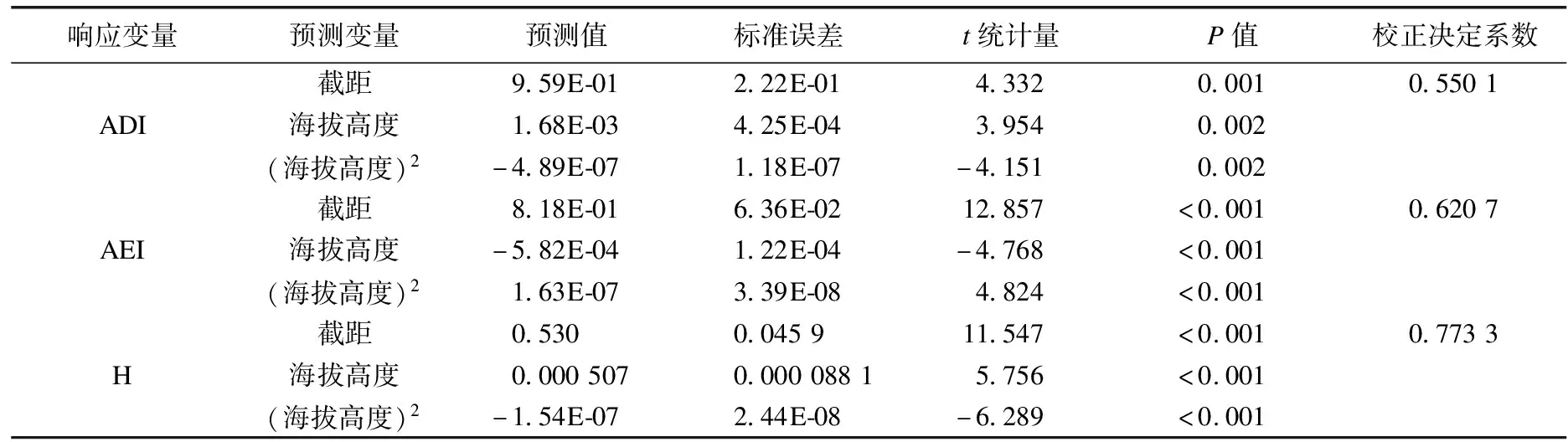
表3 回归分析结果
3 讨论
本研究发现山噪鹛声学多样性指数、声学均匀度指数、声学熵与海拔梯度存在函数关系,表明山噪鹛鸣声特征受海拔梯度影响.这一结果与云南柳莺个体鸣唱音节间隔时间与海拔存在函数关系的结果一致[16].不同在于云南柳莺个体鸣唱音节间隔时间与海拔存在正相关关系,即随海拔升高,鸣唱音节间隔时间增加;而本研究中的鸣声特征与海拔存在二次函数关系.这一差异可能是由于海拔梯度差异.云南柳莺分布海拔在1 930~3 050 m,本研究山噪鹛分布海拔为150~3 600 m.
本研究中其他声学指标与海拔梯度的函数关系不显著,声学多样性指数、声学均匀度指数、声学熵与海拔梯度关系显著.可能的原因在于,这三个指标将鸣声分成不同频率波段计算,比其他声学指标更能抓住反映鸣声的某些特征.在陈进等人进行的7个鸣声特征与鸟类多样性关系研究中,同样是声学多样性指数、声学均匀度指数、声学熵这个指标的表现优于其他指标[17].
山噪鹛的声学多样性指数与声学熵随海拔梯度增加先升高后降低,最高值出现在中海拔地区,而声学均匀度指数随海拔梯度增加先降低后升高,最低值出现在中海拔地区.声学多样性指数与声学熵指标均体现声音无序程度,无序程度越高表明声音无规律程度越高,即变化越大;声学均匀度指数体现声音的均匀程度,均匀度越低说明声音无规律程度越高.声学多样性指数、声学熵与声音均匀度指数所揭示的现象具有一致性.鸣禽具有学习同类或其他鸟类鸣声的行为[18],声音无序程度越高则表明同域分布的鸟类数量越多,即具有更丰富的鸟类物种多样性.中海拔地区鸟类物种多样性高于中、低海拔区域可能与植物物种多样性有关.垂直地带分布格局中,中海拔地区植物物种多样性高于高海拔与低海拔地区是常见格局,主要是由于降水与温度变化共同导致的[19,20].中海拔地区较高的植物物种多样性可为鸟类提供较丰富的栖息地与食物来源.
信号传达有效性是鸟类鸣声传递的重要方面[21,22].野外环境中鸟类鸣声易受噪音影响,主要体现在噪音降低声信号的传播距离、干扰声信号内容[23,24].噪音环境中鸟类往往改变声音参数如提高鸣声音量或改变鸣唱字节,加强声信号传播有效性[25].这一现象在对其它鸣禽的研究中得到证实[26].本研究中声学复杂度指数、生物声学指数、频峰数与海拔梯度不存在函数关系,表明声音复杂性未随海拔梯度发生趋势性变化.这意味着海拔梯度增加或降低,并没有改变环境中的噪音背景.
环境选择压力可塑造鸟类鸣声特征.鸣声特征不仅与背景噪音、人类活动强度等直接影响鸣声传递因素有关[27],而且与斑块大小、斑块隔离程度等生境特征[28,29]、降水等气候特征[9];纬度等水平地理特征有关[8,30].而本研究表明在垂直地理特征方面,海拔很可能是促进鸣声特征分化的重要因素.
分析动物声学特征是理解动物行为与进化的重要手段[31].鸟类鸣声分析在录音设备与分析技术的支撑下成为野外调查手段的重要补充[32].基于鸣声特征开发出一系列指标,这些指标在揭示鸣声特征方面发挥作用[33].本研究中通过声学多样性指数、声学均匀度指数、声学熵等指标揭示的海拔梯度与鸣声特征关系,表明鸣声声学特征在揭示鸟类行为与环境因素关系方面的潜力.值得指出,不同鸣声指标揭示声音特征的不同方面,并非所有指标均能实现研究目标.如生物声学指数通过计算频谱图中特定频率范围内,高于特定阈值的音量衡量群落水平的生物多样性.相比特定物种的鸣声,这一指标更适合评估群落尺度的鸣声多样性.鸣声特征研究有赖于选择适合指标.
致谢:感谢辽宁大学生命科学院万冬梅教授和殷江霞副教授提出的宝贵意见.
