程序性翻译后修饰对FEN1功能调控的研究进展
2020-06-30贾兰兰王依依华跃进
贾兰兰 王依依 华跃进 徐 虹
(浙江大学原子核农业科学研究所/农业农村部和浙江省核农学重点实验室,浙江 杭州 310029)
瓣状核酸内切酶(flap endonuclease 1,FEN1)是一种结构特异性5′核酸酶,在人类、啮齿类、两栖类、酵母、植物和古细菌中均保守存在,其主要分布于细胞核和线粒体[1],能够识别特定的核酸分叉结构,并切除含游离5′末端的单链核酸,因而得名。 除了5′-3′ flap核酸内切酶(FEN)活性[2],FEN1 蛋白还具备一定的缺口核酸内切酶(gap endonuclease,GEN)活性[3]和核酸外切酶(exonuclease,EXO)活性[4]。 研究发现,酵母中FEN1 同源基因Rad27 突变体表现出自发突变率升高的增变表型[5],并且突变体的质粒及微卫星序列表现出不稳定现象[6],Rad27 在线粒体诱变、双链断裂修复[7]、同源重组修复[8]、冈崎片段成熟[9]、线粒体DNA完整性维持[10]等过程中发挥重要作用;线虫中FEN1同源基因CRN-1 与CPS-6(Endo G)共同介导凋亡过程中DNA 的降解[3];拟南芥中,FEN1 能够维持基因组稳定性和转录基因沉默[11]。 FEN1 广泛参与细胞内的各种DNA 代谢途径,如冈崎片段成熟、长片段碱基切除修复、细胞凋亡DNA 片段化、端粒稳定性维持、停滞复制叉重启等[12-14],在正常启动DNA 复制、重组与修复[15-17],以及维持基因组的稳定性[18]等过程中发挥着关键作用。 因此,FEN1 是细胞维持遗传稳定性的重要基因之一,对FEN1 蛋白功能调控的研究具有重要的意义。
当生物体内编码FEN1 的基因出现异常时,可能会导致FEN1 在体内不正常地增多、减少或出现功能异常,从而引起基因组的不稳定,进而增加自身免疫疾病[19]、慢性炎症或肿瘤[20]的发生几率,严重危害机体健康。 研究表明,FEN1 在包括前列腺癌[21]、胰腺癌[22]、肺癌[23]、胃癌[24]、乳腺癌[25]、卵巢癌[26]、食管癌[27]、狼疮性肾炎[28]等疾病中均表达异常,导致肿瘤发生率增高或加快肿瘤的发展进程[29],已成为一种潜在的肿瘤标志物[6]。 因此,FEN1 的活性和定位必须受到极为严格的控制。
目前已知FEN1 细胞内存在3 种主要的功能调节方式:1)细胞的区室化:在正常状态下,FEN1 C 端的核定位信号基序会使其主要定位于细胞核,并高度浓缩在核仁区[30],但当细胞遭受紫外(UV)处理之后,核仁中的FEN1 会被磷酸化,并转移至核质中,通过其定位调节FEN1 的活性与功能。 2)蛋白质的相互作用:目前已鉴定出来自各种DNA 代谢途径的超过34 种蛋白质与FEN1 相互作用,如在DNA 复制的过程中,FEN1 被有效地招募到增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)上,发挥切割RNA 引物的功能。 对于长片段RNA 引物,FEN1 可以与DNA2 一起完成切割[31]。 在DNA 修复中,FEN1 能够激活长片段碱基切除修复(long-patch base excision repair,LPBER)中相关蛋白(如Polβ)酶活性,与PCNA 形成复合体发挥核心作用[32-33]。 此外,FEN1 的核酸酶活性及相互作用的Werner 综合征蛋白(Werner syndrome protein, WRN)是端粒稳定性所必需的[34],对维持基因组稳定性至关重要。 在细胞凋亡中,FEN1 与凋亡相关蛋白Endo G 的相互作用极大地增强了GEN 和EXO 活性,这对处理凋亡DNA 片段化十分重要[4]。3)翻译后修饰(post-translational modification,PTM):FEN1 可被PRMT5 甲基化(methylation)[35],被Cdk-Cyclin 激酶复合物磷酸化(phosphorylation)[36],被p300 乙酰化及发生类泛素化(SUMOylation)[37-38]和泛素化修饰(ubiquitylation)等。
近年来,蛋白质翻译后修饰研究的兴起为生物体内FEN1 功能的精细调控机制研究提供了新的思路和方向。 它们在调节FEN1 的细胞定位、特定底物酶切活性、稳定性以及蛋白质相互作用等过程中发挥着重要作用。 本文对FEN1 翻译后修饰的研究进展进行总结和分析,以期为后续研究提供参考。
1 FEN1 的翻译后修饰
1.1 磷酸化
Henneke 等[36]研究发现,在细胞S 期后期,细胞周期蛋白依赖性激酶Cdk1-Cyclin A 或Cdk2-Cyclin E复合体可以将FEN1 的丝氨酸残基(Ser187)磷酸化,该磷酸化会随着细胞周期的调节逐渐增加。 体外活性分析发现,磷酸化后的FEN1 蛋白的GEN 和EXO 活性均降低,并消除了与PCNA 的结合,但其与DNA 的结合不受影响。 此外,FEN1 突变体S187A 细胞株表现出细胞周期S 期积累,表明通过细胞周期调节的FEN1磷酸化对于复制叉的调节十分重要。 Guo 等[30]发现,FEN1 在HeLa 细胞的核仁中富集,而当细胞受到UV处理后,FEN1 会迁移到核质中并参与DNA 上的UV交联解析,在修复停滞的DNA 复制叉中发挥作用。 该过程的响应依赖于FEN1 的磷酸化修饰,磷酸化的FEN1 会协助其从核仁转移以参与响应UV 照射的DNA 损伤修复。 最新研究结果还揭示了FEN1 磷酸化在抵抗氧诱导的DNA 损伤和维持适当的细胞周期中发挥重要作用,并调节胎儿到新生儿过渡期间的心脏发育[39]。
1.2 甲基化
Singh 等[25]发现FEN1 在多种癌症中显著上调,且FEN1 的异常表达与肿瘤细胞中FEN1 启动子内的CpG 岛的低甲基化有关。 随后,Guo 等[35]鉴定了FEN1 的甲基化修饰位点,并发现蛋白质精氨酸甲基转移酶(protein arginine N-methyltransferase 5,PRMT5)能以S-腺苷甲硫氨酸(S-adenosyl methionine,SAM)作为甲基供体对FEN1 的精氨酸残基(arginine,Arg192)进行甲基化修饰。 且FEN1 的R192 位点甲基化会抑制FEN1 的S187 位点磷酸化从而确保其与PCNA 紧密结合,在DNA 复制的过程中共同发挥作用。 研究表明,赖氨酸甲基转移酶SET7 可以在体内以细胞周期依赖性方式使FEN1 的赖氨酸残基(Lys377)进行单甲基化修饰[40],K377me1 在S 期以SET7 依赖性方式上调,当离开S 期时又降低,且这一修饰对其活性没有任何影响,也未影响与重要伴侣(如BLM、PCNA 和DNA2)的相互作用。 早期研究已证明FEN1 的Lys377能够被组蛋白乙酰转移酶p300 乙酰化,以响应UV 损伤[41]。 但FEN1 K377me1 对紫外线损伤没有调节作用,也未响应甲基甲磺酸盐(mitomycin,MMS)的DNA损伤处理, 但其突变体 K377R 对羟基脲(hydroxycarbamide,HU)药物更加敏感,这表明FEN1 K377me1 位点在细胞对复制应激反应中至关重要。
1.3 类泛素化
在细胞内,FEN1 的核酸酶活性和蛋白互作需要被严格调节,与此同时,FEN1 蛋白的胞内浓度也需要被严格控制,而这一调节过程又是高度依赖于细胞周期进行的。 在S 期结束,当FEN1 完成DNA 复制后,FEN1 的去向可能对细胞的生存至关重要。 研究者提出了两种可能的假设[42]:其一,FEN1 在完成复制的复杂过程后,再次被修饰利用;其二,FEN1 在完成复杂的功能后,被降解清除,以免过度积累造成对细胞的损伤,这也是大多数蛋白质的命运。 正因如此,FEN1 的类泛素(small ubiquitin-like modifier,SUMO)化和泛素化被发现并研究。
大量研究表明,类泛素(SUMO)化在蛋白质的降解中发挥着重要作用[43-44]。 哺乳动物有4 个SUMO的旁系同源体:SUMO1、SUMO2/SUMO3 和SUMO4。SUMO 主要存在于细胞核中,其中,SUMO1 在核内广泛存在,主要修饰生理状态下的蛋白质,而SUMO2/3主要分布于核质,在应激蛋白的修饰中发挥作用,SUMO4 是新近发现的一种SUMO 分子[40],且只在肾脏、淋巴和脾脏中表达[45]。 近期,浙江大学华跃进课题组研究发现,当细胞面临UV 胁迫时,FEN1 会发生I型SUMO 化修饰[46],而且FEN1 蛋白的磷酸化作用能够促进FEN1 的I 型SUMO 化修饰,进而影响FEN1 与9-1-1 复合体的结合,参与细胞的损伤修复进程。 而FEN 1 的SUMO3 修饰则是其降解途径的一个重要步骤,FEN1 在体外或细胞内均可以发生SUMO3 修饰。质谱法鉴定出FEN1 的主要SUMO3 修饰位点位于赖氨酸残基(Lys168)。 FEN1 的突变体蛋白K168R 在体外或者细胞内均不再被SUMO3 修饰,且这一SUMO3修饰依赖于磷酸化修饰,如果FEN1 被磷酸化,则该蛋白的SUMO 化的效率会显著增加[37]。 早期研究表明,SUMO4 修饰可能会增加I 型糖尿病的患病风险[45],但在FEN1 中的功能还有待进一步研究。
1.4 泛素化
尽管SUMO 化修饰与泛素化修饰采用相似的修饰途径,且修饰过程中均有3 种酶(激活酶E1、结合酶E2 和连接酶E3)发挥作用,但修饰过程中的酶和发挥的功能却完全不同。 研究表明,FEN1 的赖氨酸残基(Lys354)会被UBE1/UBE2M/PRP19 复合物泛素化,而该过程依赖于FEN1 本身的3 型SUMO 化[37]。 被泛素化修饰后的FEN1 可以被蛋白酶体的识别,并通过泛素化酶蛋白酶体进行降解,从而调节细胞内FEN1的蛋白浓度,维持基因组的稳定。
1.5 乙酰化
Hasan 等[41]发现转录共激活因子p300 组蛋白乙酰转移酶可以将FEN1 的4 个赖氨酸残基(Lys354、Lys375、Lys377 和Lys380)乙酰化。 在该过程中,p300的HAT 结构域发挥着主要的作用,所有鉴定的乙酰化赖氨酸残基都位于C 末端,前期研究已证明FEN1 的C 末端对DNA 结合很重要[47],而且FEN1 的C 末端尾部内的大多数赖氨酸在真核生物中是保守的,说明这4 个残基的乙酰化可能是高等真核生物中进化上保守的调节机制[48]。 研究发现,FEN1 的乙酰化显著降低了FEN1 的DNA 结合能力和核酸酶活性,抑制了FEN1 与9-1-1 复合物(Rad9/Rad1/Hus1 细胞周期检查点复合物)的相互作用[49],但与PCNA 的结合能力不受影响[50]。
Wang 等[51]在进行乙酰化代谢组学分析时发现了FEN1 三个乙酰化位点(Lys80、Lys267、Lys375),但其中有两个位点(Lys80 和Lys267)在前期的体外试验中未被发现,这些新位点的功能也尚不清楚。 华跃进课题组[52]近期研究也鉴定到了FEN1 的新乙酰化修饰位点(Lys125、Lys252、Lys254 和Lys314),结果显示,这些位点的突变会影响该蛋白质的DNA 酶切活性和DNA 底物结合活性。 突变株K125A 和K252/K254A的乙酰化水平明显降低,表明这些位点是FEN1 乙酰化修饰的关键位点。 此外,细胞周期和细胞凋亡率的研究结果显示,K252A 和K252/K254A 突变株的G1期比例显著增加,K252/K254A 突变细胞对UV 诱导的DNA 损伤更敏感,凋亡细胞百分比显著增加[52],说明这些位点的乙酰化修饰在维持细胞正常生理状态和基因组稳定性中都发挥着一定的作用,但具体的作用机制仍有待进一步研究。
1.6 琥珀酰化
Weinert 等[53]对大肠杆菌(E. coli)、酵母(yeast)和人类宫颈癌细胞(HeLa cell)中的蛋白质赖氨酸乙酰化和琥珀酰化修饰组学进行了系统的分析,发现FEN1 存在(Lys200 和Lys354)琥珀酰化修饰,序列分别为HLTASEAK(su)K 和VTGSLSSAK(su)R,且这两个位点和该蛋白质鉴定到的乙酰化修饰位点是重叠的。 华跃进课题组[54]的试验结果显示,FEN1 突变体K200E 的核酸内切酶切割底物效率较FEN1 野生型(WT)降低约38%(表1),表明这一位点琥珀酰化与酶活性的相关性。 但目前有关FEN1 琥珀酰化的研究较少,关于这一位点琥珀酰化的功能也有待进一步研究。
2 FEN1 翻译后修饰的程序性调控
2.1 磷酸化和甲基化的关系
研究表明,FEN1 甲基化和磷酸化修饰会随着细胞周期的进程发生动态变化。 在DNA 复制S 期早期,FEN1 R192 的甲基化抑制了FEN1 S187 的磷酸化,使FEN1 替代聚合酶(Polδ 或Polβ)并与PCNA 发生相互作用,完成RNA 引物的切割,形成成熟的冈崎片段。非甲基化的FEN1 被Cdk/Cyclin E 复合物磷酸化,迫使FEN1 与PCNA 和DNA 底物解离,继而让连接酶LigⅠ取代FEN1 的位置完成对缺口的连接。 FEN1 的甲基化抑制其磷酸化,FEN1 的甲基化形式(而不是磷酸化形式),与增殖细胞核抗原(PCNA)强烈相互作用,确保其反应“开启”和“关闭”时间。 一旦破坏FEN1 精氨酸甲基化,其与PCNA 蛋白的相互作用也会不稳定,进而影响其定位到DNA 复制或修复位点。这两种修饰平衡的紊乱,会导致细胞内冈崎片段成熟的缺陷,细胞周期进展的延迟,DNA 损伤的累积和全基因组突变频率的升高。
2.2 磷酸化和泛素化的关系
FEN1 的泛素化是一种常见的PTM,通过将泛素分子结合到靶蛋白上,促进靶蛋白被蛋白酶体识别,诱导其降解。 而SUMO 化是一种不同的PTM,不介导靶蛋白的蛋白酶体降解,而是能够可逆性的修饰靶蛋白,进而参与其定位及功能调控[42]。 FEN1 的类泛素化和泛素化共同在FEN1 的降解中发挥作用。 首先,FEN1的磷酸化会促进其发生类泛素化,FEN1 的类泛素化修饰诱导FEN1 的泛素化修饰,使得泛素化修饰的FEN1 在蛋白酶体的介导下降解。 FEN1 的SUMO 化在FEN1 蛋白的降解过程中起着类似于信号的作用,诱导其继续发生泛素化修饰,继而准确地调节细胞内FEN1 的活性和表达量[42]。
2.3 乙酰化和琥珀酰化的关系
FEN1 的乙酰化和琥珀酰化修饰均发生于赖氨酸残基上。 赖氨酸乙酰化被认为是在细胞生理学和病理学等方面的重要调节机制;赖氨酸琥珀酰化是一种新发现的类似于赖氨酸乙酰化的翻译后修饰类型,在各种生物体中广泛存在。 由于赖氨酸琥珀酰化比乙酰化的结构更为复杂,这种修饰便可能诱导FEN1 的化学性质及其结构与功能发生更大的变化[55]。 赖氨酸乙酰化和琥珀酰化是蛋白质酰基化的主要类型,在许多细胞过程中都是重要的,包括基因转录、细胞代谢、DNA 损伤响应等。 FEN1 琥珀酰化和乙酰化修饰是动态调节的,在不同细胞和组织中存在修饰位点差异。此外,赖氨酸乙酰化和赖氨酸琥珀酰化的修饰位点的重叠暗示这两种修饰之间可能存在着某种互补或者拮抗作用。 然而,FEN1 蛋白的这两种修饰在细胞中的具体生物学功能和调节方式仍有待进一步挖掘。
近十几年,FEN1 蛋白翻译后修饰的研究发现,该蛋白质在细胞内存在着多种翻译后修饰类型。 这些修饰或相互协作,或相互拮抗,影响着FEN1 的细胞定位、酶切活性、蛋白互作及蛋白质本身的稳定性。 在细胞周期S 期早期,FEN1 发生甲基化修饰,甲基化的FEN1 取代Polδ 与PCNA 密切结合,在分枝结构处进行切割。 当FEN1 被去甲基化后,继而会发生磷酸化修饰,迫使FEN1 从PCNA 和DNA 底物的复合体上解离,继而连接酶LigⅠ取代FEN1 的位置完成对缺口的连接[29]。 磷酸化的FEN1 在细胞内存在两种去向[56],第一种是细胞处于正常生长状态时,在S 末期从复制复合物上脱落下来,作为FEN1 的降解起始步骤,促进FEN1 的2/3 型SUMO 化,继而诱导泛素化蛋白酶降解体降解;第二种是当细胞在复制过程中遇到了一定的外界胁迫,导致细胞内出现了大量的DNA 损伤,此时,磷酸化的FEN1 则会被1 型SUMO 化,从而更易于被9-1-1 复合体招募到损伤的DNA 位点,进行修复(图1)。 这些修饰过程环环相扣,高度协调,一旦其中某种修饰无法正常产生,则会导致细胞周期的紊乱,错误DNA 的积累,甚至还会造成染色体的异常分离,进而影响细胞的基因组稳定性。 此外,FEN1 的乙酰化修饰和Ⅰ型SUMO 化、磷酸化一样,也会在UV 等DNA损伤剂处理下有所提升[41],然而其在DNA 损伤响应和修复中的生物学意义尚未知。 这三种修饰(乙酰化、磷酸化、SUMO 化)之间是否有一定的联系仍不清楚。
3 总结与展望
据不完全统计,研究者们已鉴定出的蛋白质的翻译后修饰种类已经超过400 种。 几乎所有的蛋白质均可发生PTM,且同一种蛋白质还可能同时发生多种PTM,共同有序地调节着细胞的生命活动。 PTM 的发生会改变蛋白质的理化性质、结构与功能,每一个小的PTM 可能显著改变蛋白质的活性与功能,而多种PTM的共同发生,又将产生极其复杂的变化。 植物中,琥珀酰化会改变蛋白的溶解性等性质,在重金属、高盐高温、辐照等胁迫条件下,农作物的DNA 的甲基化会产生显著差异,严重威胁到农作物生长[57-58]。 总之,同一种蛋白质,即使只发生一种PTM,也可能赋予不同的功能;发生在不同氨基酸上的同一种修饰也将产生不同的功能状态;不同种修饰的功能及参与不同生物过程就更加复杂。 PTM 种类的复杂性及其对蛋白质功能调控的多样性共同决定了这是一个具有挑战性的研究领域。
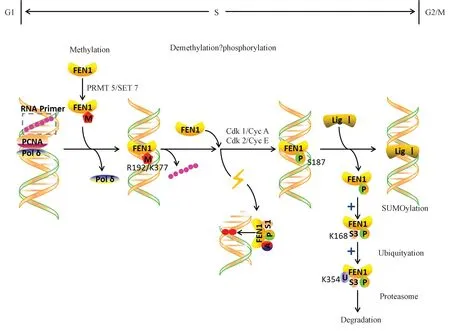
图1 FEN1 在细胞周期中程序性翻译后修饰的研究模型[29,31]Fig.1 FEN1 model for post-translational posttranslational modification in cell cycle[29,31]
近十几年来,有关FEN1 翻译后修饰的研究也已经取得了相当大的进展。 研究者建立了FEN1 翻译后修饰的周期依赖程序性修饰模型,并勾勒出主要的框架,但针对整个翻译后修饰组学来讲,这些研究仅是冰山一角。 如,早期研究已经证明FEN1 C 末端的几个氨基酸残基(K354、K375、K377 和K380)能够被组蛋白乙酰转移酶p300 乙酰化以响应UV 损伤[41]。 同时,这些乙酰化修饰位点还存在着其他的修饰类型,如K354 还可以发生琥珀酰化, K377 位点还能被甲基化等。 但这些位点发生不同类型修饰的具体生理状态和所需条件,以及不同修饰后FEN1 蛋白的具体生物学功能仍需进一步深入研究。
由于FEN1 在不同物种中的高度保守性,使得这些基础研究有较高的借鉴性。 因此,如何通过调控FEN1 蛋白的功能或表达量来维持基因组的稳定性,进而提高物种的抗逆性,是未来研究的一个重要方向,该方向,包括FEN1 蛋白的翻译后修饰改变与物种特性变化的相关性关系,以及FEN1 蛋白的乙酰化和琥珀酰化在植物代谢中的作用等。
在FEN1 蛋白自身复杂的翻译后修饰调控的同时,FEN1 的互作蛋白的修饰也可能存在着变化。 因此,要明确这一过程的生物学意义,仅局限于FEN1 蛋白的修饰显然不够。 例如,p300 除了能使FEN1 乙酰化,还能使DNA2 乙酰化;乙酰化的DNA2 能以更高的亲和力结合其DNA 底物,并刺激其5′-3′核酸内切酶、5′-3′解旋酶和DNA 依赖性ATP 酶活性[59]。 p300 对这两种内切核酸酶活性的差异调节表明,乙酰化能够促进细胞中较长片段形成的机制,从而更有效地解析冈崎片段成熟和DNA 损伤修复过程中产生的片段结构。 此外,Takawa 等[60]发现组蛋白赖氨酸甲基转移酶SETD8 可以在PCNA 的赖氨酸残基(Lys248)上进行甲基化修饰,PCNA 和FEN1 之间的不同亲和力并不受FEN1 的磷酸化状态的调节,而是受PCNA 的甲基化状态的调节,表明这些修饰将可能存在着更为复杂的调节机制。 因此,结合细胞生物学和结构生物学研究动态的蛋白质修饰组学十分必要。
