特色油料作物油莎豆CeDGAT1基因的鉴定及表达分析
2020-06-19王晓清高慧玲薛金爱李润植
高 宇,陈 莹,孙 岩,王晓清,高慧玲,薛金爱,李润植
(山西农业大学分子农业与生物能源研究所,山西太谷030801)
莎草科莎草属植物油莎豆(Cyperus esculentus)是多年生草本植物(野生型),其栽培种为1年生春播作物。油莎豆分蘖力强,须根发达,须根顶端可膨大为圆形或椭圆形的地下块茎。与马铃薯等地下块茎类作物不同,油莎豆地下块茎含油量可达35%(干质量)以上,且产量和出油率较高,抗逆性及适应性强,管理简单[1]。油莎豆油营养价值与橄榄油相当,其抗氧化性强,且健脾健胃,可预防心血管病以及高血脂等心血管系统疾病,是优质的保健食用油[2]。同时,油莎豆块茎油脂也可作工业用油和化工产品原料油,其制备高品质生物柴油的最高转化率可达94.5%以上[3]。因此,研究油莎豆块茎油脂合成和积累机制,以及培育高产、高油新种质,将有助于这种特色油料作物产业链提质增效和可持续发展。
三酰甘油酯(TAG)是油料作物油脂的主要储存形式,主要于内质网上合成[4]。二酰甘油酰基转移酶(Diacylglycerol acyltransferase,DGAT)负责使用酰基-CoA底物在1,2-二酰甘油(DAG)的sn-3位置进行酰化,生成TAG[5],该酶促反应是TAG合成的限速步骤之一,并且在控制油脂积累量和油脂成分方面发挥重要作用[6]。迄今,植物中已经被鉴定出4种类型的DGAT,即DGAT1、DGAT2、水溶性的DGAT3[7]和双功能蜡酯合酶WS/DGAT[8]。已有研究表明,大豆(Glycinemax)、蒺藜苜(Medicagotruncatula)、旱金莲(Tropaeolum majus)等植物的不同组织中都检测到DGAT1基因的表达,尤其是种子中的表达量较高,且表达量同TAG积累趋势成正比[9]。拟南芥(Arabidopsis thaliana)中过量表达AtDGAT1,会导致种子千粒质量和含油量都明显增加[10]。在拟南芥的ASII突变体中,DGAT1活性降低,含油量下降超过20%[11]。将DGAT1-2基因转入高油玉米(Zea mays)中,其转基因籽粒含油率提高6.19%[12]。与DGAT1基因相比,DGAT2基因在有特异脂肪酸合成的植物中对TAG合成的作用更大,二者之间可能无功能冗余[13]。DGAT3基因编码的酶蛋白是无跨膜区,存在于细胞质的水溶性蛋白,明显区别于膜结合的DGAT1和DGAT2家族,且起源不同、独立进化[14]。现今,DGAT1基因已经在不同的植物中被克隆和研究,包括拟南芥、烟草(Nicotiana tabacum)、甘蓝型油菜(Brassica napus)、蓖麻(Ricinus communis)、大豆、油桐树(Vernicia fordii)等。研究显示,DGAT1对油脂合成和积累具有重要的贡献和影响[10,15-17]。在我们建立的油莎豆块茎转录组数据中,发现了一个编码DGAT1的cDNA(CeDGAT1)序列,可能在块茎油脂生物合成积累中行使重要功能。
本研究通过生物信息学分析油莎豆CeDGAT1的理化性质、功能结构、跨膜结构域、二级结构、系统发育,以及通过实时荧光定量检测CeDGAT1在油莎豆块茎不同发育时期的表达谱,进而鉴定CeDGAT1在油莎豆块茎油脂及脂肪酸合成途径中的功能,旨在为提高油莎豆块茎油脂产量的遗传改良和解析油脂合成代谢调控机制提供科学依据。
1 材料和方法
1.1 供试油莎豆种质资源
试验所需的油莎豆种质材料为山西农业大学分子农业与生物能源研究所保存的晋农1号,种植于山西农业大学农作站。油莎豆CeDGAT1的编码序列来源于山西农业大学分子农业与生物能源研究所实验室的转录组数据。
1.2 油莎豆CeDGAT1编码蛋白序列的生物信息学分析
用NCBI中的Conserved Domain Database(CDD)库(http://www.ncbi.nlm.nih.gov/Strucdd/cdd.shtml)鉴定CeDGAT1蛋白的保守结构域;通过MEME(http://meme-suite.org/)预测CeDGAT1蛋白的Motif;利用Expasy网站提供的Protparam工具分析油莎豆CeDGAT1蛋白的氨基酸组成、相对分子质量、理论等电点和原子组成等理化性质;使用一系列在线工具分析CeDGAT1蛋白跨膜区(http://www.cbs.dtu.dk/services/TMHMM/)、疏水区域(http://web.expasy.org/protscale/)和信号肽(http://www.cbs.dtu.dk/Services/SignalP/);使用PBIL LYON-GE-RLAND和Phyre 2对CeDGAT1蛋白的二级结构和三级结构进行预测分析;使用Genedoc软件对油莎豆CeDGAT1和拟南芥AtDGAT1进行氨基酸的多序列比对分析;使用MEGA 7.0软件采用邻接法(Neighbor-Joining,NJ)对油莎豆CeDGAT1和其他物种的DGAT1蛋白序列(表1)构建系统发育树,自举检验值为1 000个循环,其余值均为默认设置。

表1 不同植物DGAT1氨基酸序列信息
1.3 油莎豆CeDGAT1基因的表达检测
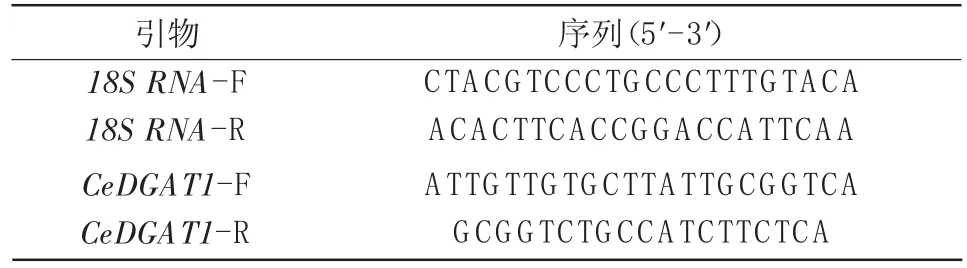
表2 定量PCR引物
采用北京全式金公司的Trans ZolPlant试剂盒提取油莎豆发育块茎的总RNA;使用Genstar公司的反转录试剂盒将提取的RNA反转录为cDNA;利用Primer 6.0设计CeDGAT1序列的qRT-PCR引物,以18SRNA为内参基因(表2)。荧光定量采用Takara的反应体系:定量正向和反向引物各0.4μL、SYBRRPremix Ex TaqⅡ(2×)5μL、ROX Reference Dye(50×)0.2μL、RNase-free H2O 3μL、cDNA模板1μL。反应程序为:95℃10 min;95℃15 s,58℃1 min,40个循环,5次生物学重复。
2 结果与分析
2.1 油莎豆CeDGAT1蛋白的理化性质分析
利用CDD分析油莎豆CeDGAT1蛋白的功能结构,与拟南芥相比,油莎豆CeDGAT1蛋白与拟南芥AtDGAT1蛋白都属于MBOAT超家族,但是CeDGAT1没有PLN02401功能区域(图1);Motif预测结果显示,CeDGAT1与AtDGAT1结构相似,都包括4个相同的Motif(图2)。对油莎豆CeDGAT1蛋白的理化性质进行分析和鉴定,结果显示(表3),CeDGAT1编码502个氨基酸,分子质量为57.68ku,预测理论等电点为9.23;预测的不稳定系数为53.33,为不稳定蛋白;该蛋白包括8个跨膜结构域;脂肪族系数和亲水系数表明,该蛋白为疏水性蛋白。CeDGAT1编码的氨基酸数较AtDGAT1少,但是二者都是疏水性的跨膜蛋白。

表3 油莎豆CeDGAT1和拟南芥AtDGAT1蛋白的理化性质分析
2.2 油莎豆CeDGAT1蛋白的高级结构预测分析
油莎豆CeDGAT1蛋白是由α-螺旋、延伸链、β-折叠、无规则卷曲组成(图3),其中,α-螺旋占比最高,为46.06%;其次是无规则卷曲,占比为35.43%;延伸链占比为12.40%;β折叠占比最低,为6.10%,与拟南芥AtDGAT1中二级结构元件组成相同、结构相似。通过Phyre 2在线软件对At-DGAT1和CeDGAT1进行同源建模,预测其三级结构,结果显示(图4),以c6buhH一种膜蛋白为模板,AtDGAT1和CeDGAT1蛋白与模板的相似性都为58%,二者的三级结构极其相似。
2.3 油莎豆CeDGAT1蛋白的序列比对及系统进化树分析
使用Genedoc对油莎豆CeDGAT1和拟南芥AtDGAT1的序列比对结果显示(图5),CeDGAT1和AtDGAT1的序列相似性为61%;对比其他DGAT1蛋白的保守序列,CeDGAT1的氨基酸序列中包含有预测的DGAT保守序列:酰基-CoA结合位点、催化活性位点、磷酸泛素结合位点等;在其预测的巯基辅酶位点的序列中包含一个保守的脯氨酸,脯氨酸被认为参与脂肪酸酰基基团的传递[18];预测的脂肪酸结合蛋白位点包括一个酪氨酸磷酸化位点;预测的DAG/佛波酯结合序列包含一个保守的苯丙氨酸位点。油莎豆CeDGAT1存在多个功能位点,预示其可能具有DGAT酶活性。
CeDGAT1与一些已知的DGAT1序列构建系统进化树,结果显示(图6),CeDGAT1与单子叶植物高粱SbDGAT1和野生水稻ObDGAT1的亲缘关系比较近;而与拟南芥、蓖麻、花生、大豆的亲缘关系较远,分别分布于2个分支中。
2.4 油莎豆CeDGAT1在油莎豆不同发育时期块茎中的表达分析
为了检测油莎豆CeDGAT1在块茎发育过程中的表达情况,通过荧光定量PCR(qRT-PCR)检测CeDGAT1在块茎不同发育时期的表达,并以18SRNA为内参基因,计算不同发育时期的表达情况,结果显示(图7),CeDGAT1在块茎发育的不同时期均可表达,相对表达量先上升后下降,且在块茎播种90 d时的表达量最高,此阶段也是块茎油脂快速富集的时期。
3 结论与讨论
为确保粮油安全和经济可持续发展,特别是为解决由于植物油用于制备生物柴油而对全球粮食安全的威胁,开发产油量高的新型植物油脂资源和进一步提高现有油料作物的产量,为近几年研究的一个热点领域。地下营养器官块茎富油的油莎豆,产量也高于现有商业化的大田油料作物,其成为人们聚焦研究的一个新型特色油料作物。DGAT在油脂合成中具有重要作用,使得其成为人们研究的重点。油莎豆自20世纪被我国引进以后,大部分研究偏向油脂提取工艺对油品质的影响、生物柴油制备、组分分析、栽培技术等方面,关于油莎豆油脂代谢机制以及提高油脂含量代谢工程的研究日渐备受重视[19-20]。
本研究对油莎豆的CeDGAT1基因功能进行了逐步鉴定,结果显示,油莎豆CeDAGT1基因编码502个氨基酸,是不稳定的疏水性蛋白,有8个跨膜结构,是与内质网膜结合的跨膜蛋白。功能结构域分析显示,油莎豆CeDGAT1蛋白属于MBOAT超家族,其二级结构和三级结构显示出与拟南芥AtDGAT1极大的相似性。多序列比对结果显示,CeDAGT1的氨基酸序列包含DGAT典型的保守区、酰基转移酶结合位点和催化活性位点,其中,DAG/佛波酯结合位点是DGAT特有的[21]。进化树分析结果显示,油莎豆CeDGAT1与单子叶植物高粱、野生水稻亲缘关系较近,推测CeDGAT1具有DGAT酶活性。
油莎豆CeDGAT1基因在块茎发育的所有时期均有表达,在块茎播种90 d时表达量达到最高,表达趋势呈单峰曲线。现在已经报道的不同植物的DGAT1,有不同的组织和时空特异性表达水平。花生AhDGAT1-1在花和种子中的表达水平高于其他组织,而AhDGAT1-2在根、种子和子叶中显示出了更高的表达水平[22]。拟南芥AtDAGT1基因在大部分的组织中都有表达,但在发育中的胚胎和花瓣中表达量最高[23];紫苏PfDGAT1在不同组织中均有表达,且在种子中表达量最高[24]。据报道,在烟草和酵母中过量表达拟南芥AtDAGT1基因大大提高了转化品系的TAG含量[10,25]。在蓖麻中,RcDGAT1在叶片和发育中的种子之间几乎没有差异,并且RcDGAT1在10 DAP(授粉天数)的表达最高[26]。在斑鸠菊中,VgDGAT1的表达水平在胚中远高于根、茎、叶和果皮。在种子发育期间,VgDGAT1表达水平从早期(10~17 DAP)缓慢增加,然后在24 DAP时急剧上升到最大值,随后VgDGAT1表达量逐渐下降[27]。油莎豆发育中块茎的油脂含量增加分为含油量低、增加缓慢的早期以及油脂快熟积累的中期和含油量高增长速度逐渐平缓的后期[28],这种增长趋势与CeDAGT1的表达模式相似,因而推测,CeDGAT1在油莎豆块茎油脂积累中具有重要的作用。
通过对油莎豆块茎油脂中关键基因的发掘和验证,对于解析营养器官大量积累油脂的机制具有重要的作用。本研究结果可为后期试验中利用分子育种和基因工程等手段改良油莎豆品质以及提高油莎豆油含量和油产量提供重要理论依据,也将为油莎豆分子育种提供新思路。
