模拟穿透雨减少对锐齿栎(Quercus aliena var. acuteserrata)树干液流密度的影响
2020-06-12凌海燕刘世荣栾军伟刘晓静牛晓栋张京磊
凌海燕,刘世荣,,*,栾军伟,刘晓静,牛晓栋,张京磊,李 想
1 国际竹藤中心竹藤资源与环境研究所, 北京 100102 2 中国林业科学研究院森林生态环境与保护研究所, 北京 100091 3 河南宝天曼国家级自然保护区管理局, 南阳 474350
伴随着全球气候变化,区域的降水格局发生了显著改变,造成水分分配不均和季节性干旱的现象[1]。这不仅极大地影响了区域水循环,而且还影响了土壤-植被-大气连续体系统[2]。森林生态系统具有极强的碳固持和水源涵养功能,树木的蒸腾耗水对区域水循环和碳循环均有较大的影响,因此,植被蒸腾成为了全球气候变化生态学研究关注的重要问题。树干液流密度可以长期、连续和较准确地反映单木水分蒸腾[3]。通过测定树干液流密度,探求树木的蒸腾耗水特征及其对环境变化的响应,在森林生态系统的水分循环研究中得到广泛应用[4- 5]。
不同树种的树干液流密度在不同环境条件下呈现出不同的变化特征,大多数研究只是定性地描述了树种之间的差异[6]。同一树种(如:毛白杨Populustomentosa、雪松Cedrusdeodara、香樟Cinnamomumcamphora)的整树蒸腾量随着胸径的增大而增大[7]。树木胸径、边材面积[8]、太阳辐射以及空气水汽压亏缺等[9]均可影响树木的蒸腾量。较大的树木因其根系较深和茎干储水能力强,能更好地适应夏季干旱[10]。然而,红桦(Betulaalbosinensis)的蒸腾作用与土壤含水量之间无直接关系[11]。连续2个月的短期减雨处理对菜豆树(Radermacherasinica)和圆叶乌桕(Sapiumrotundifolium)的树干液流密度无显著影响[12]。美国西部科罗拉多州的美洲山杨(Populustremuloides)[13]和澳大利亚东北部昆士兰州的桉树(Eucalyptusrobusta)[14],由于干旱,造成树木水分供应不足,发生了大面积死亡的现象。在长白山移除2个月的林内穿透雨后,土壤含水量下降了24.6%,但成熟蒙古栎(Quercusmongolica)的光合生理、水分生理并没有受到影响[15]。以树干液流密度为代表的树木水分代谢过程对干旱响应的生理生态学机制还缺乏深入的研究。
锐齿栎(Quercusalienavar.acuteserrata)作为优势树种广泛分布于华北暖温带落叶阔叶林区,在河南宝天曼地区常形成以锐齿栎为单优势种的落叶阔叶林[16]。本研究在宝天曼森林生态站建立模拟干旱的野外固定实验样地,利用热扩散探针技术,测定锐齿栎不同径级树干液流密度的动态变化,分析其对模拟林内穿透雨减少后的响应规律及其原因。研究目的是:(1)揭示模拟降雨减少下锐齿栎树干液流密度的季节变化规律,探究锐齿树干液流密度对干旱的响应;(2)不同径级锐齿栎的树干液流密度对干旱的响应特征是否一致;(3)阐明干旱胁迫环境条件下,树干液流密度与叶片生理参数及环境因子之间的关系。在此基础上,评价锐齿栎对气候变化背景下干旱的响应,解析干旱对锐齿栎水分利用特征的影响。本研究可为认识锐齿栎林生态系统的耗水特征及其对气候变化背景下干旱的响应提供基础数据,也为制定暖温带落叶阔叶林适应气候变化的管理对策提供科学依据。
1 实验地概况和研究方法
1.1 研究区域
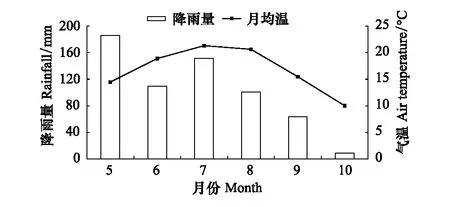
图1 2018年生长季降雨及气温变化特征Fig.1 Characteristics of rainfall and air temperature change in the growth season of 2018
研究地点位于河南省西南部的南阳市内乡县宝天曼国家级自然保护区内的宝天曼森林生态系统定位研究站,属伏牛山南坡(111°47′—12°04′ E,33°20′—33°36′ N),是暖温带向亚热带过渡地区,属大陆性季风气候。宝天曼的最高海拔为1830 m,研究地海拔约1400 m。年均降水量900 mm,年蒸发量991.6 mm,年平均气温11—14 ℃,气温在年际间的变化比较稳定。2018年观测期内5月份降雨量最高,为186.2 mm,5—8月份降雨量均超过100 mm,10月份降雨量只有8.8 mm;最高气温为7月份,月均温达21.3 ℃,生长季内雨热基本同期(图1)。研究地区植物种类丰富,共有种子植物150科783属2147种,植被区划属暖温带落叶阔叶林,林冠层优势树种是锐齿栎,占林分树种组成的65%,林分平均年龄约为65年,锐齿栎的树高基本在20 m左右,2015年锐齿栎的平均胸径为25.2 cm[16]。样地内亚冠层小乔木伴生种有短柄枹(Quercusbrevipetiolata)、四照花(Dendrobenthamiajaponica)、化香(Platycaryastrobilacea)、三桠乌药(Linderaobtusiloba)、水榆花楸(Sorbusalnifolia)等。
1.2 减雨实验设计
依托宝天曼森林生态系统定位研究站,在以锐齿栎为优势树种的天然林中,建立了6块(20×20 m2)标准样地,设计了林内穿透雨减少(减雨)和对照2种不同的实验处理,每个处理3个重复。在减雨样地内安装1.5—2.5 m高的不锈钢钢架搭上减雨膜(主要拦截林内穿透雨),由导水槽将截留的雨水导出样地。每块减雨样地四周均设有70 cm深的透明隔水板,防止样地内外土壤水分交换。样地建于2013年,每年(5—10月)对减雨样地进行覆膜减雨,减雨膜的覆盖面积占样地面积的50%,减雨量约占年降雨量的30%[17]。2018年5月开始将减雨膜覆盖面积增加至70%,实际样地减雨量约占年降雨量的40%,以适度增加减雨样地的水分胁迫。
2018年4月在每块减雨样地和对照样地中分别选取15棵生长良好,树干通直、无被挤压现象的锐齿栎作为测定树干液流密度的对象。每个样地锐齿栎分三个径级(胸径≤21 cm为小径级、21—30 cm为中径级、≥30 cm为大径级),每个径级长期连续测定5棵。由于2018年7月遭受强雷击事件,造成部分观测仪器设备损坏,因此,锐齿栎树干液流密度的数据只采用了一个减雨样地和一个对照样地在2018年5—10月份的数据。减雨样地和对照样地样树的平均胸径分别为25.5、26.3 cm,平均冠幅分别为14.2、16.7 m2,边材面积分别为188.9、198.1 cm2,样树基本特征见表1(对照样地实际有效样树只有13棵)。
1.3 测定指标及方法
1.3.1树干液流密度的测定
2018年4月对选定锐齿栎安装探针,Granier探针安装方法参照赵平等[3],即在锐齿栎树干胸高处、正北方向进行安装。将探针与数据采集器CR1000-ST(Campbell Scientific, Logan, UT, USA)和通道扩展版AM16/32B(Campbell Scientific, Logan, UT, USA)相连,每10 s读一次数据,每10 min进行平均并记录一次数据,每隔一星期导取数据并使用软件计算树干液流密度。
探针读取的原始电压值,需要借助Granier经验公式换算为液流密度值[18]:
式中,(Js)为瞬时液流密度(sap flux density)(g m-2s-1),(ΔTm)为昼夜最大温差,(ΔT)为瞬时温差。瞬时液流密度(Js)的计算采用美国杜克大学环境学院植物生理生态实学验室Ram Oren团队开发的Basliner 3.0.8软件[10]。

表1 样树的基本特征
TR: 减雨Throughfall reduction;CK: 对照 Control
1.3.2环境因子测定
(1)气象因子数据来源于宝天曼国家森林生态系统定位研究站的蛮子庄自动气象站,降雨量(Rainfall)由翻斗式自计雨量计(Davis Rain collector II model 7F852, Davis Instruments, USA)测定、光合有效辐射(Photosynthetically Active Radiation)由(Model LI190SB, Li-cor, Inc., USA)测定、空气温度(Air Temperature)和空气相对湿度(Air Relative Humidity)由(Model HMP45D Vaisala, Finlan)测定。将每半小时的平均值自动记录于数据采集器CR3000(Campbell Scientific, Logan, UT, USA)和通道扩展模块AM16/32B(Campbell Scientific, Logan, UT, USA)内。
(2)每个减雨样地与对照样地内均在距土壤表面5 cm、20 cm、30 cm设有5TM传感器(Decagon Devices, Inc., USA),5TM传感器可以测定土壤温度(Soil Temperature)和土壤含水量(Soil Moisture Content),数据每半个小时平均一次并记录,由Em50 Series数据采集系统(Decagon Devices, Inc., USA)进行储存。
1.3.3植物叶片光合生理的测定
使用便携式光合仪(LI- 6400XT,Li-Cor,Lincoln Inc.NE,USA)测定锐齿栎叶片的光合速率(Photosynthetic Rate)、气孔导度(Stomatal Conductance)以及蒸腾速率(Transpiration Rate),由于锐齿栎树高基本在20 m左右,直接冠层测量存在一定困难,因此采用离体测量法。每个样地用高枝剪剪下向阳的枝条放入水中每次测定3个叶片,每个叶片重复测3次,2018年5—9月每月测定1次日变化(7:00—18:00)。
1.4 数据处理
采用SPSS 20.0软件进行独立样本T检验,分析减雨样地和对照样地不同径级锐齿栎树干液流密度的差异。采用Pearson相关分析,研究液流密度和光合速率、气孔导度以及蒸腾速率的相关性,分析影响锐齿栎树干液流密度的叶片光合生理参数。采用逐步多元回归分析,研究液流密度和空气温度、光合有效辐射、空气相对湿度、土壤温度和土壤含水量的关系,分析树干液流密度的驱动因子在降雨减少后的变化。
2 结果与分析
2.1 穿透雨减少对生长季内土壤温度和土壤含水量的影响
2018年生长季内,不同深度的土壤温度随时间均呈先升高后下降的趋势(图2),5—8月份减雨样地与对照样地均表现为表层土壤温度(5 cm)高于深层土壤温度(30 cm),9—10月份则表现为表层土壤温度(5 cm)低于深层土壤温度(30 cm)。不同深度的土壤含水量变化与降雨事件存在很大关系。对照样地的土壤含水量在5—8月份表现为:30 cm>20 cm>5 cm,在9—10月份则表现为:30 cm>5 cm>20 cm。减雨样地不同深度的土壤含水量变化较大,整体表现为减雨样地的土壤含水量小于对照样地。

图2 生长季样地土壤温度和土壤含水量动态Fig.2 Dynamics of soil temperature and soil moisture content in the growth seasonTR: 减雨Throughfall reduction, 即,TR5、TR20、TR30分别表示减雨样地距土壤表面5 cm、20 cm、30 cm的土层深度数据; CK: 对照 Control, 即:CK5、CK20、CK30分别表示对照样地距土壤表面5 cm、20 cm、30 cm的土层深度数据
2.2 穿透雨减少对锐齿栎树干液流密度的影响
在生长季内,减雨样地和对照样地中不同径级的锐齿栎树干液流密度在不同月份的月均值(忽略夜间液流密度,取每天5:00—19:00的数据求得日均值再进行月均值计算)有所不同(图3),6、7月份减雨样地中的平均树干液流密度小于对照样地,而随着减雨处理时间的累积,8—10月份减雨样地的平均树干液流密度大于对照样地。小径级的树干液流密度,在5—7月份减雨样地低于对照样地,而在8—10月份减雨样地高于对照样地;且小径级的树干液流密度在5、9、10月份有显著性差异,在7月份达到极显著性差异。中径级的树干液流密度,只在7月份减雨样地显著低于对照样地;且在5、7、10月份有极显著性差异。大径级的树干液流密度,在6—9月份减雨样地低于对照样地;且在7、10月份有极显著性差异,在6月份有显著性差异。

图3 不同径级的树干液流密度Fig.3 Js of different diameter classesTR: 减雨Throughfall reduction;CK: 对照 Control; **表示处理间液流密度差异极显著(P<0.01);*表示处理间液流密度差异显著(P<0.05)
2.3 生长季内树干液流密度与叶片生理参数及环境因子的相关关系
2.3.1锐齿栎的树干液流密度与叶片生理参数的相关关系
从表2可以看出,在减雨样地,锐齿栎树干液流密度与气孔导度及蒸腾速率存在极显著的正相关关系(P<0.01),与光合速率存在显著的正相关关系(P<0.05);在对照样地,锐齿栎树干液流密度与光合速率、气孔导度、蒸腾速率均存在极显著的正相关关系(P<0.01)。
2.3.2锐齿栎的树干液流密度与环境因子的关系
对不同径级锐齿栎的树干液流密度与环境因子进行逐步多元回归分析表明(表3),锐齿栎的树干液流密度均受空气相对湿度和光合有效辐射的影响,锐齿栎树干液流密度与空气相对湿度呈负相关,与光合有效辐射呈正相关。对照样地锐齿栎树干液流密度的影响因子有土壤含水量,而减雨样地中土壤含水量这个因子不进入回归模型。建立的回归模型,均具有显著性,判定系数R2也较大,因此模型能较好地说明不同径级的锐齿栎树干液流密度随影响因子变化而变化的规律。

表2 液流密度与光合速率、气孔导度、蒸腾速率的Pearson相关关系
**表示极显著性相关(P<0.01); *表示显著性相关(P<0.05)
3 讨论
由图1和图2可以看出,生长季内降雨量少的月份,土壤含水量也较少,减雨样地的土壤含水量显著小于对照样地。林内穿透雨减少对锐齿栎树干液流密度产生了影响,随着时间累积,减雨样地的树干液流密度显著高于对照样地(图3)。Fisher等[19]在亚马逊河东部的实验发现经减雨处理会降低林分蒸腾量,Besson等[20]通过在葡萄牙南部进行的截留降雨和灌溉试验同样得出截留降雨后降低了林分蒸腾量的结论。而锐齿栎树干液流密度在干旱条件下上升的原因,由于随着减雨处理时间加长,土壤含水量降低,促使锐齿栎呈现适应性响应,吸收更多水分来维持树木本身水分的需求。以往的研究结果也有发现,对于一些落叶树树种,干旱并不会导致其树干液流密度下降,但叶片会在生长季的后期提前衰老、凋落[21]。锐齿栎为落叶阔叶树种,其生长季与雨季对应,因此,锐齿栎生长季内极少出现严重或长期的缺水干旱现象;而降水较少的春、冬季节与锐齿栎非生长季对应,其对水分的需求较低,因此不影响锐齿栎生长。干旱在一定程度上可以提高植物的水分利用效率,土壤含水量对干旱时期林分蒸腾有显著影响[9],且对液流密度的总体水平起到决定作用[22]。锐齿栎通过在干旱时提高树干液流密度以满足树干和叶片的水分需求。

表3 液流密度与环境因子的多元线性回归
TR: 减雨Throughfall reduction, CK: 对照Control;Ta: 空气温度Air temperature,RH: 空气相对湿度 Air relative humidity,PAR: 光合有效辐射Photosynthetically active radiation,Ts: 土壤温度Soil temperature,SWC: 土壤含水量 Soil water content
减雨样地与对照样地不同径级的锐齿栎树干液流密度存在差异,小径级的树干液流密度经减雨处理后,受到的影响比其他径级大,说明胸径大小在一定程度上影响树干液流密度与土壤含水量的关系(图2、图3),与熊伟等[23]研究华北落叶松(Larixprincipis-rupprechtii)得出的结论一致。罗超等[24]发现土壤表层含水量降低时,地下水会在毛管力的作用下使土壤表层水分得到补充,以满足杨树(Populus)的生长需求。小径级的锐齿栎冠幅较小,相对应的根系也较小,易受浅层土壤含水量的影响,吕金林等[25]研究辽东栎(Quercusliaotungensis)发现小径级的树木同样对土壤含水量变化敏感。万艳芳等[26]研究发现,青海云杉(Piceacrassifolia)优势木与被压木相比具有较大的平均液流密度,原因是具有较大冠幅和胸径的优势木能获得更多的水分、营养等资源。可能是由于胸径较大的树木存在较厚的导水及储水组织,使树干液流密度不易受土壤含水量的影响。
无论在减雨样地还是在对照样地,锐齿栎树干液流密度跟叶片光合速率、气孔导度、蒸腾速率均存在显著正相关关系(表2)。叶片水平上的光合速率和蒸腾速率与单株水平上的树干液流密度虽呈显著性相关,但仍表现出一定的不同步[21]。气候变化后,植物会在生理水平上进行一定的调整,短时间内通过调整叶片气孔导度来适应环境变化[27]。叶片气孔导度的大小决定了树干液流密度的大小,气孔可以根据环境条件的变化来调节自己开度的大小而使植物在损失水分较少的条件下获取最多的CO2[28],从而得到最大的光合有效速率。陈志成等[16]在同一实验样地进行锐齿栎的水力结构研究发现,穿透雨减少后,锐齿栎的叶片气孔密度变大,但气孔长度变小,说明了减雨样地的锐齿栎发生了适应性调节。
树干液流密度在时间尺度上的变化主要是由各种环境因子的共同作用造成的(表3)。姚依强等[29]发现树干液流密度与环境因子的关系会随时间而发生显著改变。本研究中,减雨样地与对照样地不同径级的锐齿栎树干液流密度均与光合有效辐射呈正相关关系,而与空气相对湿度呈负相关关系,这与卢志朋等[30]研究结果一致。宝天曼具有极大的空气相对湿度(生长季的大部分时候达90%以上),保证了锐齿栎生长时的水分需求,即使加大对林内穿透雨的截留面积,周围空气中的水分也会扩散到减雨样地。在降雨减少后,减雨样地与对照样地的所有环境驱动因子中最大的不同就是土壤含水量。土壤含水量的适度减少会提高植物的水分利用效率[31],但本研究的结果表明,土壤含水量不是影响减雨样地树干液流密度的重要因子。
4 结论
在暖温带锐齿栎天然林中通过穿透雨减少模拟气候干旱,土壤含水量显著降低,导致锐齿栎树干液流密度呈现干旱胁迫响应。锐齿栎树干液流密度在生长季6—7月份对林内穿透雨减少处理后呈现下降趋势,但在生长季的5月份和8—9月份不同径级锐齿栎出现差异性响应,特别是小径级的锐齿栎液流密度对土壤含水量下降响应多变,表明不同径级锐齿栎对水分胁迫呈现不同的适应性响应。在以后的研究中,还需要长期和深入地加强穿透雨减少处理后锐齿栎树干液流密度与叶片冠层生理、树木地上茎干和地下根系生长等相互作用关系的研究。
