亚东鲑幼鱼饲料蛋白和脂肪适宜水平的研究
2020-06-12高擘为梁高阳李小勤冷向军
高擘为 杨 航 何 明 徐 禛 梁高阳 李小勤 冷向军
(1. 上海海洋大学水产科学国家级教学示范中心, 上海 201306; 2. 上海海洋大学农业部鱼类营养与环境生态研究中心, 上海201306; 3. 水产动物遗传育种中心上海市协同创新中心, 上海 201306)
亚东鲑, 学名为褐鳟(Salmo trutta), 原产于欧洲、非洲北部和西亚地区[1], 属于鲑形目(Salmoniformes)鲑科(Salmonidae)鲑亚科(Salmoninae)。身体为褐色, 并有黑色及红色的斑点, 栖息在寒冷的淡水中。在我国, 褐鳟仅天然分布于西藏亚东县,是英国人于1866年引种于此, 在当地形成的一个自然种群, 已定名为亚东鲑[2], 西藏自治区二级重点保护水生动物[3]。亚东鲑肉质细嫩鲜美、营养丰富,氨基酸种类齐全, 具有优质的脂肪酸[4]。随着亚东鲑养殖业的发展, 与其相关的养殖、遗传和生物学研究的工作已相继开展[5—8]。
脂肪和蛋白质的需要量是鱼类营养研究中的重要参数。若饲料中蛋白质含量不足, 则鱼体生长缓慢, 若蛋白质过量, 成本增加, 还会造成饲料的浪费。目前, 有关褐鳟蛋白质营养需求的研究较少。Arzel等[9]的研究表明, 体重为1.2 g的褐鳟对饲料蛋白质需要量在48%—53%; Ramezani[10]发现, 体重为7.0 g的褐鳟对蛋白的最适需要量为50%。而关于褐鳟脂肪营养的研究则主要集中在不同脂肪源的替代方面, Kenari等[11]发现用植物油部分替代鱼油可长期喂食里海褐鳟。Amiri等[12]也认为大豆油和鹅油可以完全替代鱼油, 对褐鳟生长性能和存活率没有负面影响。国内关于亚东鲑鱼营养需求的研究则仅见亲鱼对饲料蛋白质和脂肪需求方面, 当饲料脂肪水平为9%和18%时, 亚东鲑亲鱼对蛋白质需要量分别为43.11%和45.69%[13]。在亚东鲑幼鱼的营养需求方面, 目前尚未见相关报道。由于亚东鲑已形成独立的种群, 以及亚东当地自然条件的特殊性(海拔高、水温低), 故开发适合于当地条件的亚东鲑饲料显得尤为必要。故本试验设计不同蛋白和脂肪水平的饲料, 考察对亚东鲑鱼幼鱼生长、饲料利用、体组成、肝脏生化指标和胃、肠道消化酶活性的影响, 为亚东鲑幼鱼配合饲料的开发提供依据。
1 材料与方法
1.1 试验设计与试验饲料
本试验采用3×2双因素完全随机设计, 设3个蛋白质水平(P, 42%、46%、50%)和2个脂肪水平(L,12%、16%), 共配制6种饲料 (P42/L12、P42/L16、P46/L12、P46/L16、P50/L12、P50/L16)。通过调整沸石粉和纤维素的含量来平衡各组饲料组成。饲料原料经粉碎、过筛(60目)后, 用混合机(GH 200, 上海展望机电设备有限公司)混匀, 制成粒径2 mm的沉性颗粒饲料(单螺杆挤压机, SLP-45, 中国水产科学研究院渔业机械仪器研究所, 制粒温度85—90℃), 晾干, 密封储藏于阴凉干燥处备用。饲料原料购于浙江粤海饲料有限公司。试验饲料的配方组成及营养成分含量见表1, 饲料氨基酸组成见表2。
1.2 试验鱼和饲养管理
试验用鱼取自西藏亚东鲑鱼繁育基地。试验鱼用基础饲料暂养驯化2周后, 选取体格健壮、规格均一, 平均体重为(2.80±0.10) g的鲑450尾进行试验, 将鱼随机分配到18个塑料桶(直径0.55 m, 高0.65 m)中, 每个试验组三个重复(桶), 每桶25尾。养殖用水为经过滤的溪水, 流水养殖, 水流量为1.8 L/min。试验期间, 每天投喂两次(10:30、17:30), 日投饲率约为鱼体重的2.0%—3.0%, 各桶保持基本一致的投饲量, 并根据摄食情况进行调整。养殖期间,水体溶氧>6 mg/L, pH 7.5—8.0, 水温7.5—12.5℃,亚硝酸盐浓度<0. 0.05 mg/L, 氨氮浓度<0. 1 mg/L。养殖试验在西藏亚东县鲑鱼繁育基地(春丕)进行,养殖周期为56d。
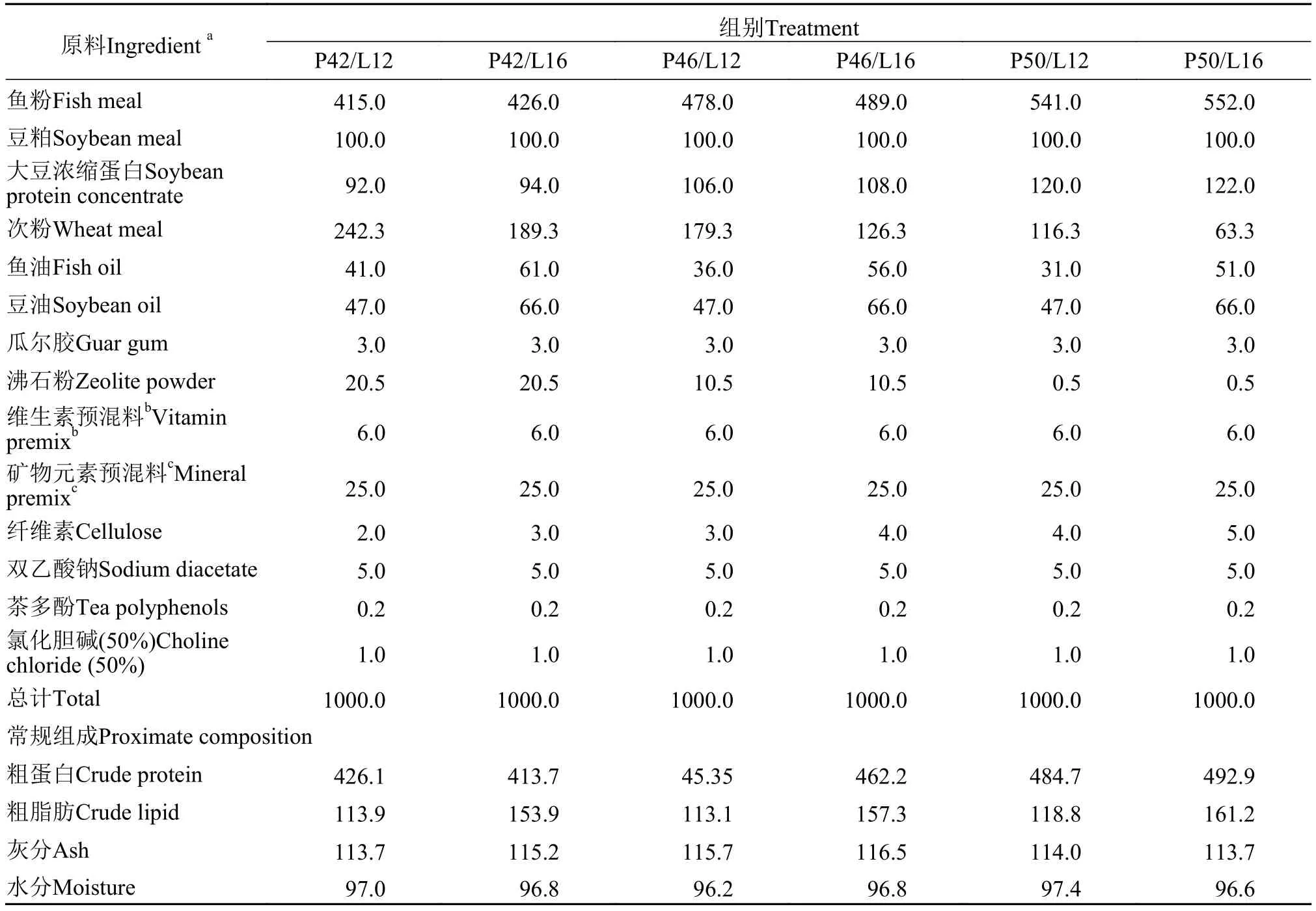
表1 试验饲料配方组成及营养成分含量Tab. 1 Ingredients and proximate composition of experimental diets (g/kg)
1.3 样品采集
在养殖试验结束后, 饥饿24h, 统计每桶亚东鲑鱼尾数并称重。从每桶中随机取6尾, 并在-20℃冷冻保存用于全鱼体成分分析。每桶随机取3尾鲑鱼测量体重和体长, 然后将鲑鱼解剖, 称量内脏和肝脏重, 计算肥满度(CF)、脏体比(HSI)和肝体比(VSI)。另取3尾鱼的肝脏、胃和前肠, -20℃冷冻保存, 用于肝脏生化指标和胃、肠消化酶活性测定。
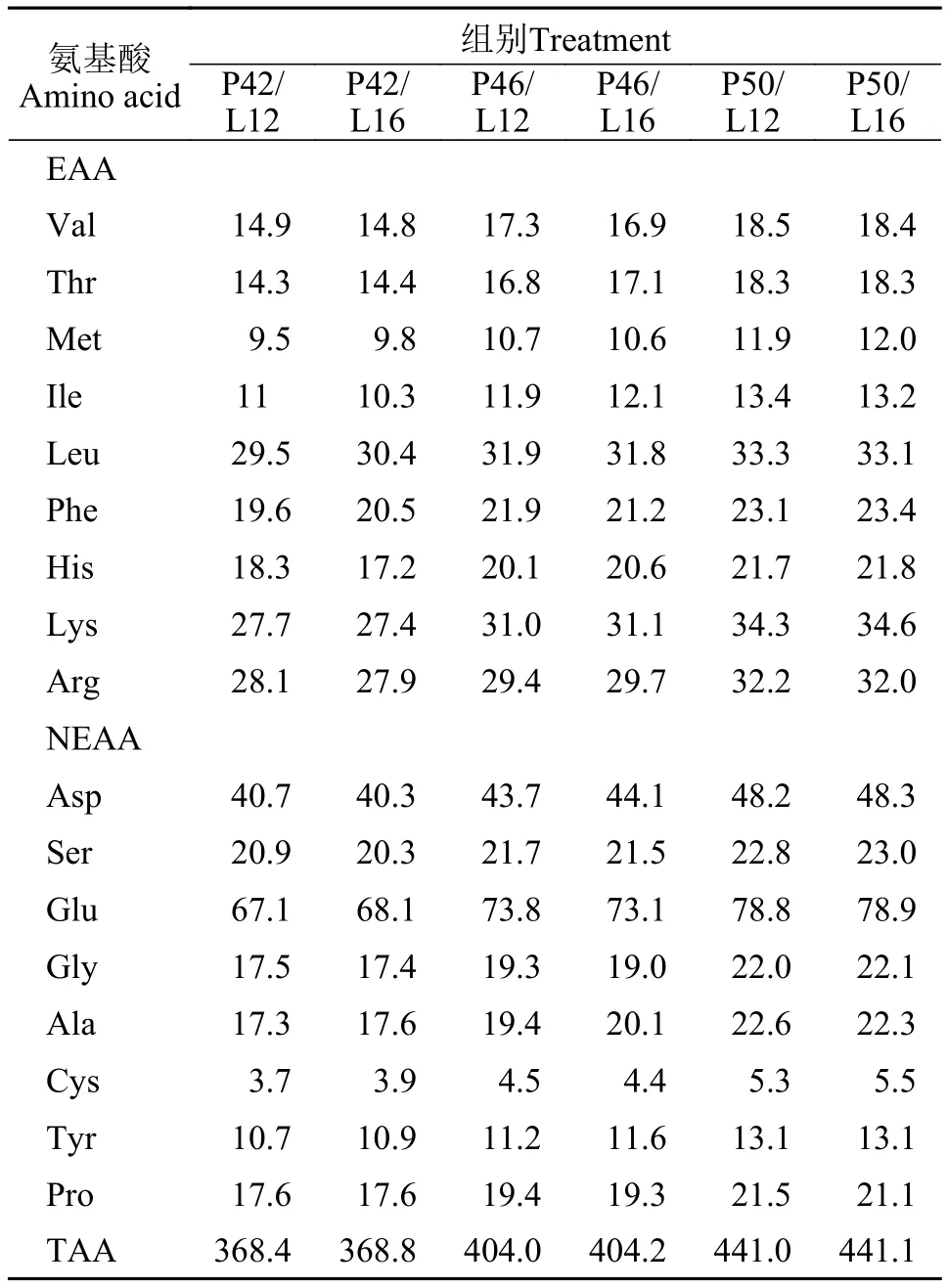
表2 试验饲料氨基酸组成(g/kg干物质)Tab. 2 Amino acid composition of experimental diets (g/kg dry matter)
1.4 测定指标与方法
生长性能与形体指标增重率(%)=100×[终末体重(g)-初始体重(g)]/初始体重(g);
饲料系数=采食量(g)/[终末体重(g)-初始体重(g)];
成活率(%)=100×终末尾数(尾)/初始尾数(尾);
肥满度(g/cm3)=100×体重(g)/体长(cm)3;
肝体比(%)=100×肝脏重(g)/体重(g);
脏体比(%)=100×内脏重(g)/体重(g);
蛋白质沉积率(%)=100×鱼体蛋白质贮积量/摄入的蛋白质总量;
脂肪沉积率(%)=100×鱼体脂肪贮积量/摄入的脂肪总量。
肝脏生化指标和胃肠道消化酶活性测定取肝脏样本于4℃解冻, 加9倍体积生理盐水, 冰水浴匀浆, 4℃离心(3000 r/min, 10min), 取上清液用于测定肝脏谷草转氨酶(GOT)、谷丙转氨酶(GPT)、总胆固醇(TCHO)和甘油三酯(TG)。采用南京建成生物工程研究所试剂盒测定GOT、GPT、TCHO和TG。
取胃、肠道样本于4℃解冻, 加9倍体积生理盐水, 冰水浴匀浆, 4℃离心(3000 r/min, 10min), 取上清液用于测定胃和肠道的蛋白酶及淀粉酶活性。肠道蛋白酶活性采用福林酚法测定, 以2%酪蛋白溶液为底物, 每微克组织蛋白质在pH 7.2、37℃条件下每分钟分解酪蛋白生成1 μg酪氨酸的酶量为1个蛋白酶活性单位(U)。胃蛋白酶采用南京建成生物工程研究所提供的试剂盒测定, 每毫克组织蛋白37℃每分钟分解蛋白生成1 μg氨基酸相当于1个蛋白酶活力单位(U)。蛋白质含量的测定采用考马斯亮蓝法。淀粉酶活性采用南京建成生物工程研究所提供的试剂盒测定, 组织中每毫克蛋白质在37℃与底物作用30min, 水解10 mg淀粉定义为1个淀粉酶活性单位(U)。
全鱼与饲料组成全鱼与饲料的水分、粗蛋白含量测定分别采用105℃常压干燥法和凯氏定氮法(2300自动凯氏定氮仪, FOSS, 瑞典), 粗脂肪含量测定采用氯仿-甲醇抽提法, 粗灰分测定采用550℃高温灼烧法。
取70 mg冷冻干燥后的饲料样品, 以6 mol/L盐酸(加1 g/L苯酚)于110℃水解24h(真空状态), 冷却,取0.5 mL水解液, 烘干, 加5 mL稀释液, 使用Sykam S-433D氨基酸自动分析仪(赛卡姆,德国)测定饲料氨基酸组成。
1.5 数据处理
试验数据以平均数±标准差表示, 采用SPSS 24.0软件进行单因子方差分析和双因素方差分析, 其中差异显著者进行Duncan多重比较, 差异显著水平为P<0.05。
2 结果
2.1 生长性能和形体指标
由表3可见, 随饲料蛋白水平的增加, 增重率呈现先上升后下降的趋势, 饲料系数则先下降后上升(P<0.05)。P46/L12组的鱼体增重率最高, 饲料系数最低; P50L12组的肥满度显著高于其他各组(P<0.05); 肝体比随着蛋白水平的增加呈现降低的趋势;各组脏体比无显著差异(P>0.05)。饲料蛋白质和脂肪水平及二者交互作用均显著影响鱼体增重率和饲料系数(P<0.05), 饲料中蛋白质和脂肪水平对亚东鲑鱼肥满度和脏体比均无显著性影响(P>0.05)。饲料中蛋白水平对亚东鲑鱼肝体比具有显著影响(P<0.05), 但两因素之间并无显著交互作用(P>0.05)。
2.2 全鱼组成和营养物质沉积率
由表4可见, 各组在全鱼水分、粗脂肪、粗蛋白和粗灰分含量上均无显著性差异(P>0.05)。饲料蛋白质和脂肪水平以二者交互作用均显著影响鱼体蛋白质沉积率和脂肪沉积率(P<0.05), 在同一脂肪水平下二者呈现先上升后下降的趋势。
2.3 胃、肠道消化酶活性
由表5可见, 饲料中脂肪水平对肠道蛋白酶, 胃蛋白酶和胃淀粉酶活性均有显著性影响(P<0.05),饲料蛋白水平仅对胃蛋白酶活性具有显著性影响(P<0.05)。两者的交互对肠道淀粉酶、胃蛋白酶和胃淀粉酶具有显著性影响(P<0.05)。46%蛋白水平下的胃蛋白酶活性显著高于其他2个蛋白水平下的胃蛋白酶活性(P<0.05), 12%脂肪水平下的胃蛋白酶、胃淀粉酶活性也显著高16%脂肪水平下的胃蛋白酶、胃淀粉酶活性(P<0.05)。
2.4 肝脏生化指标
由表6可见, 在同一脂肪水平下, GPT、TCHO和TG随着饲料粗蛋白水平的增加呈现先上升后下降的趋势。TCHO在P46L12组达到最高值, 且显著高于其他各组(P<0.05)。饲料粗蛋白水平对各肝脏生化指标均有显著性影响(P<0.05); 饲料粗脂肪水平仅显著影响TCHO含量(P<0.05)。
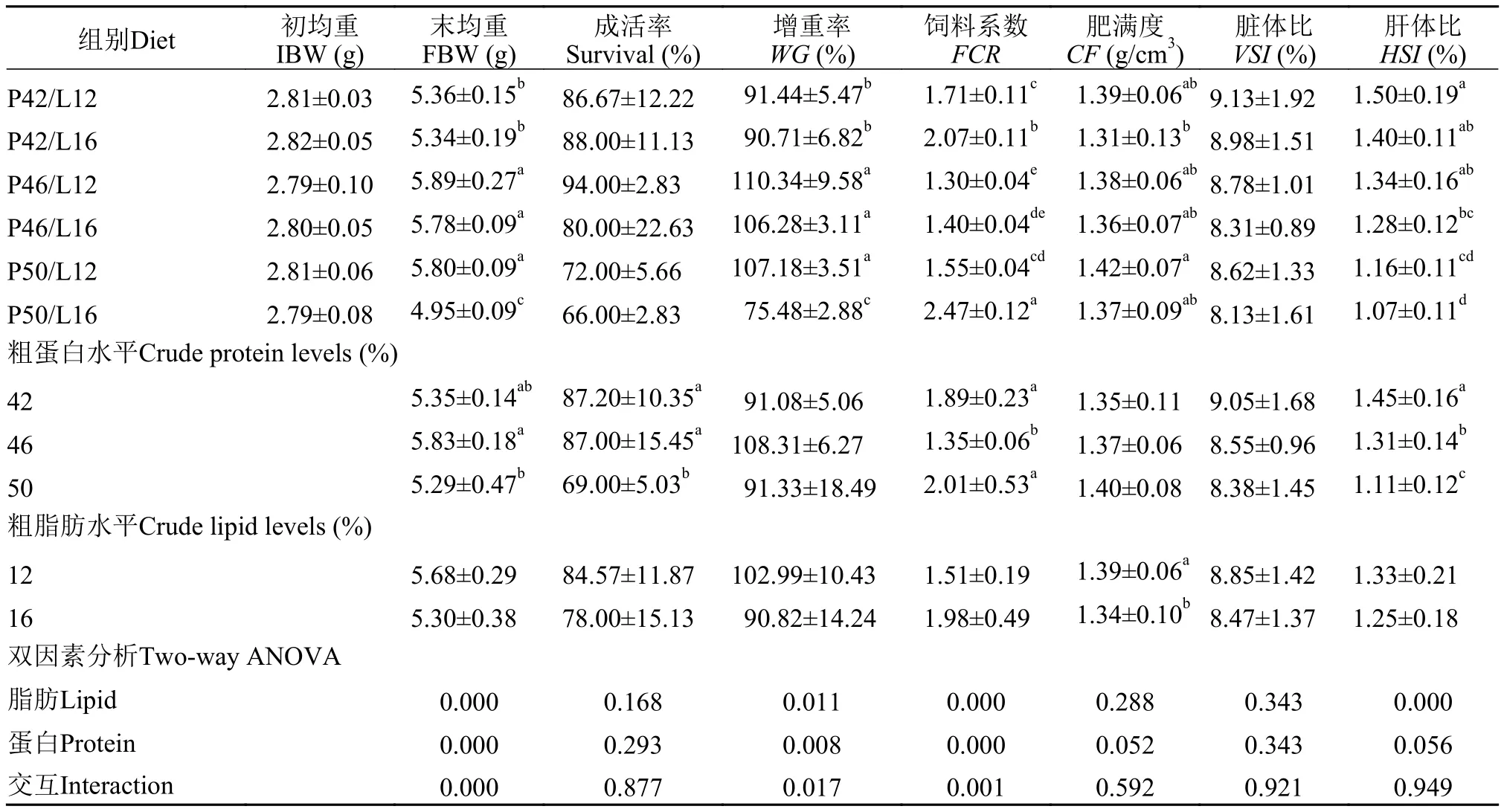
表3 饲料粗蛋白、粗脂肪水平对亚东鲑鱼生长性能的影响Tab. 3 Effects of dietary crude protein and crude lipid levels on growth of brown trout
3 讨论
3.1 饲料蛋白和脂肪水平对亚东鲑幼鱼生长性能及全鱼组成的影响
在本实验中, 在同一脂肪水平下随着饲料蛋白水平提高, 亚东鲑幼鱼增重率呈现先升高后降低的趋势, 饲料系数呈现先降低后升高的趋势。在黄斑蓝子鱼(Siganus canaliculatus)[14]的研究中, 饲料蛋白水平从24%增加到40%, 幼鱼增重率也呈现先升高后降低的趋势。同样, 在南方鲇(silurus meridionalis)[15]和淡水石斑鱼(Cichlasoma managuense)[16]中也有类似报道。可见, 鱼类对饲料蛋白质的需求具有适宜水平, 过高或过低, 均不利用鱼类生长和饲料利用。在本实验中, 亚东鲑幼鱼的饲料蛋白质需要量为46%。在Arzel等[9]和Ramezani[10]对褐鳟的研究中, 所确定的饲料粗蛋白适宜水平分别为48%—53%及50%。本试验所得结果与之相比略低,其原因与鱼体规格大小、褐鳟品系(淡水褐鳟vs.海水褐鳟)和水温等因素有关。本试验所用褐鳟属淡水种, 未经选育, 规格也较Arzel等[9]所用褐鳟(1.2 g)大。有研究表明, 鱼类的摄食和饲料利用受到水温的影响, 水温较高可促进鱼类的消化吸收[17]。在本实验中, 水温较低, 亚东鲑生长较慢, 过高的饲料蛋白可能被用作能量, 影响了亚东鲑的生长和饲料利用。
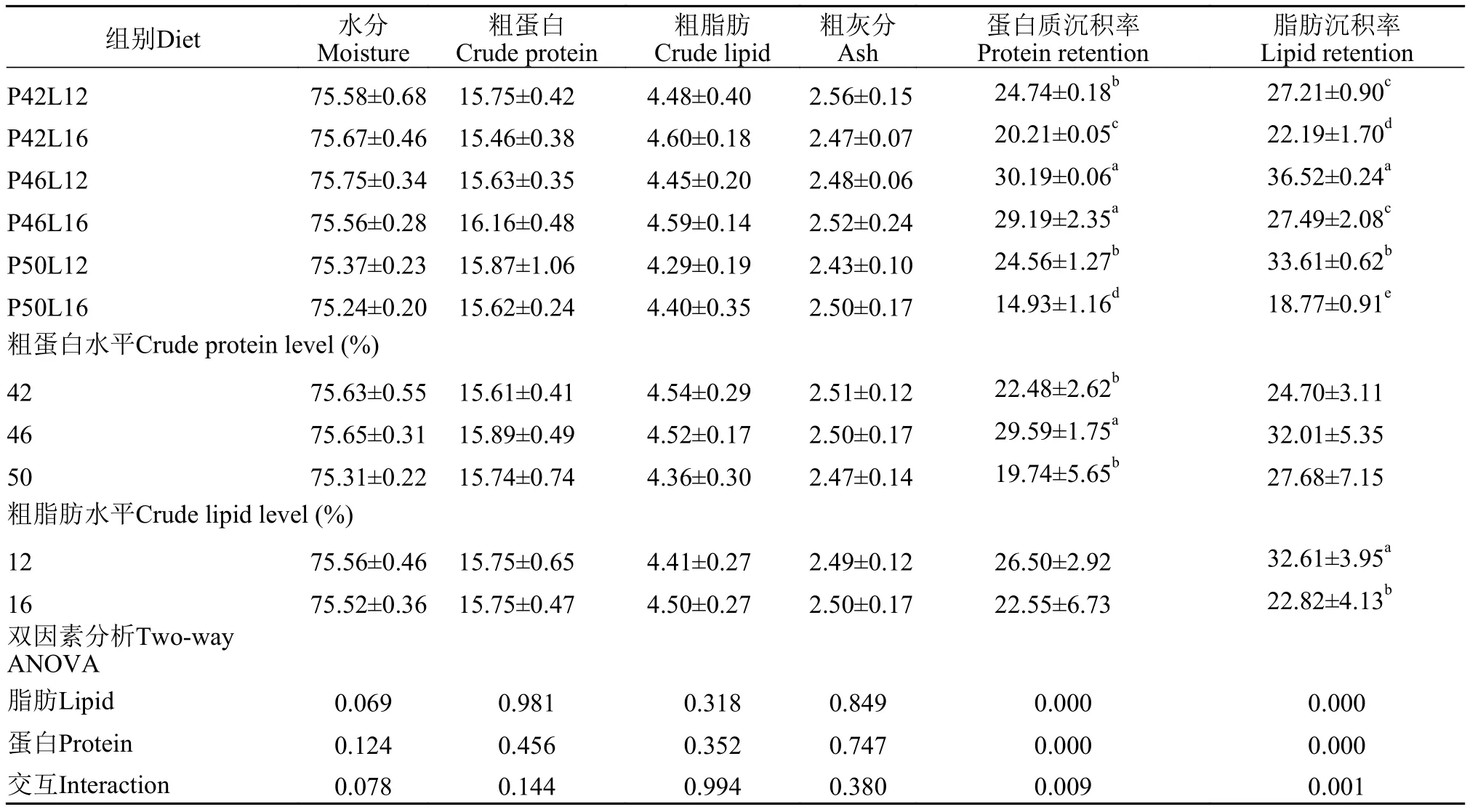
表4 饲料粗蛋白、粗脂肪水平对亚东鲑鱼全鱼组成和营养物质沉积率的影响Tab. 4 Effects of dietary crude protein and crude lipid levels on body composition and nutrient retention of brown trout (%)
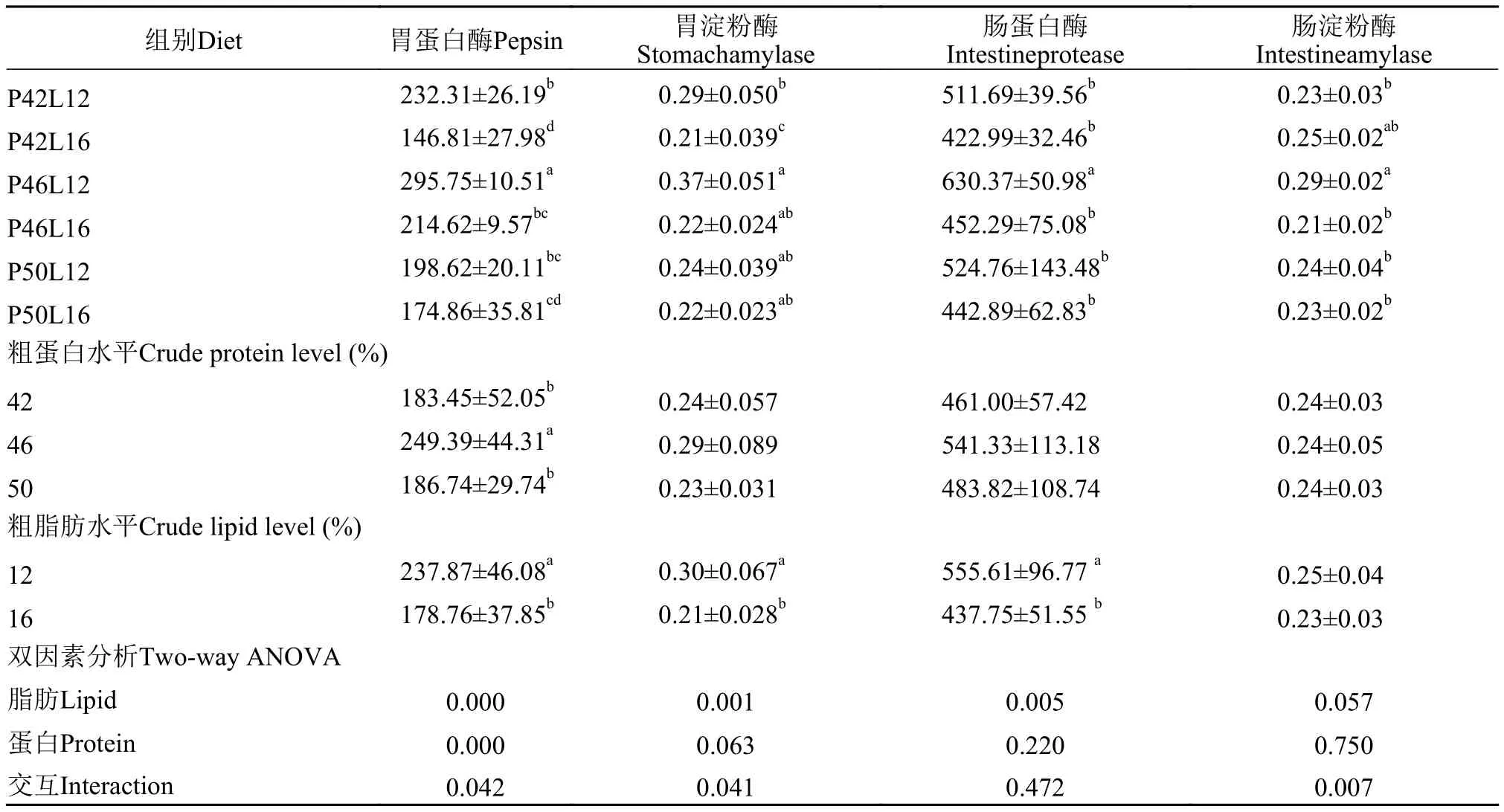
表5 饲料粗蛋白、粗脂肪水平对亚东鲑鱼消化酶活性的影响Tab. 5 Effects of dietary crude protein and crude lipid levels on digestive enzyme activities of brown trout (U/mg prot)
研究发现, 虹鳟(Oncorhynchus mykiss)[18]和南方鲇[19]的饲料脂肪添加量达20%和15%, 表明肉食性鱼类具有很强的脂肪代谢的能力, 进而有效节约了蛋白添加量。在本研究中, 当饲料蛋白质水平在42%和46%时, 脂肪水平从12%提高到16%, 尽管褐鳟增重率在数值上有下降, 但并不显著(P>0.05); 而蛋白质水平达到50%时, 16%脂肪组的增重率较12%脂肪组显著降低(P<0.05)。这说明高蛋白高脂肪饲料造成了饲料利用的降低和生产性能的下降。López等[20]发现, 当饲料蛋白水平为61%时, 饲料脂肪水平从15.5%增加到21.5%, 锤形石首鱼 (Atractoscion nobilis)的增重从21.5 g下降到14.7 g。在红鳍东方鲀(Takifugu rubripes)[21]和细鳞鲑(Brachymystax lenok)[22]的研究中, 均发现在高饲料蛋白水平下, 饲料脂肪含量增加导致了生长性能的下降。根据本试验结果, 亚东鲑幼鱼可能并不需要过高脂肪营养即可满足生长需求, 其饲料脂肪需要量为12%, 与6.9 g的虹鳟脂肪需求量(8%—10.6%)相比略高[23]。
鱼类的营养状态可由肝体比和脏体比等形态学指标反映出来。在本研究中, 饲料蛋白和脂肪水平对亚东鲑幼鱼脏体比无显著影响, 而肝体比随着蛋白和脂肪水平的提高, 呈现出逐渐下降的趋势,同样在草鱼(Ctenopharyngodon idella)[24]、欧洲鲈(Dicentrarchus labrax)[25]和黄姑鱼(Nibea albiflora)[26]的研究中也发现肝体比随着饲料中脂肪的增加而下降的现象。
在本试验中, 各组亚东鲑幼鱼全鱼水分、蛋白质、灰分和脂肪含量无显著差异。且蛋白、脂肪和两者交互作用对全鱼水分、蛋白、灰分和脂肪均无显著性影响(P>0.05)。蛋白质沉积率和脂肪沉积率随着饲料蛋白水平的升高表现出先增加后下降的趋势, P46L12组的蛋白沉积率和脂肪沉积率最高。在叶文娟等[27]的研究中, 随着蛋白水平的增加,泥鳅(Misgurnus anguillicaudatus)的蛋白沉积率也呈现先上升后下降的趋势, 说明过多的蛋白质被当作能量物质消耗, 从而降低了蛋白质沉积效率。
3.2 饲料蛋白和脂肪水平对亚东鲑幼鱼消化酶的影响
消化酶对摄食和相关代谢调整的反应可用于评估饲料配方的营养价值。在本试验中, 脂肪因素对于肠道蛋白酶, 胃蛋白酶和胃淀粉酶均具有显著性影响, 表现为在同一蛋白水平下, 脂肪升高, 消化酶活性降低。在王朝明等[28]关于胭脂鱼(Myxocyprinus asiaticus)的研究中, 脂肪水平从2.04%增加到13.39%, 蛋白酶活性也呈现下降的趋势。其原因可能是因为过高的脂肪含量抑制了消化酶的活性。
在本研究中, 在同一脂肪水平上, 随着饲料蛋白水平的升高, 消化酶活性呈现先上升后下降的趋势。这说明在适宜蛋白水平内, 亚东鲑幼鱼可通过提高体内消化酶的活性来适应饲料, 以提高对饲料的消化与吸收能力。在芙蓉鲤鲫(Carassius auratus gibelio)[29]、瓦氏黄颡鱼(Pelteobagrus vachelli)[30]、刺鲃(Barbudes caldwell)[31]和方正银鲫(Carassius auratus gibelioBloch)[32]的研究中也有类似结果。高饲料蛋白组(50%)的肠蛋白酶和胃蛋白酶活性降低,表明过量的蛋白质造成了浪费, 加重了消化道的负担, 影响了肠道消化酶的分泌[33]。淀粉酶出现先上升后下降的趋势, 可能因为饲料中蛋白和脂肪水平的变化导致碳水化合物含量的变化, 引起淀粉酶活性的变化。
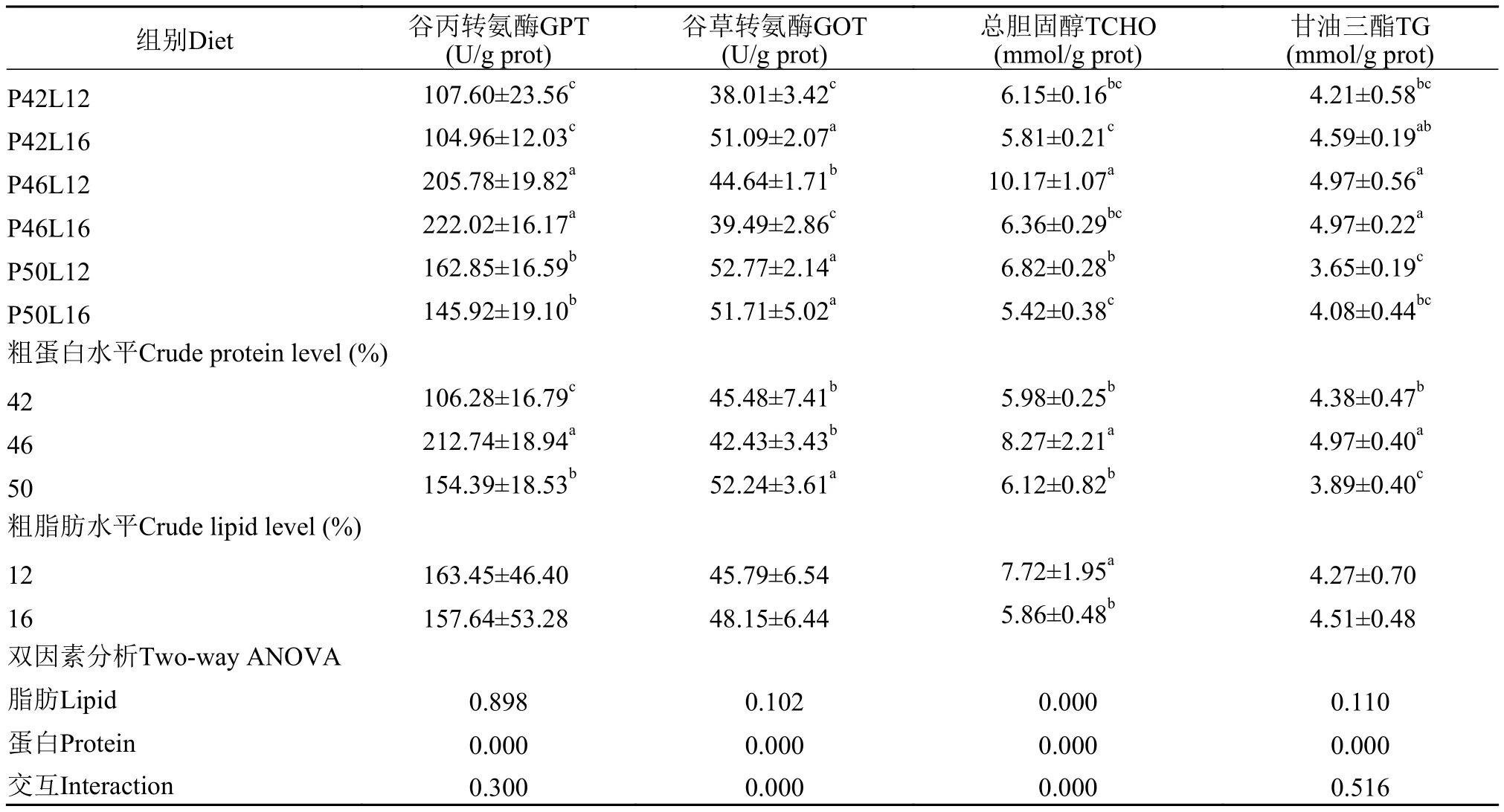
表6 饲料粗蛋白、粗脂肪水平对亚东鲑鱼肝脏生化指标的影响Tab. 6 Effects of dietary crude protein and crude lipid levels on liver biochemical indices of brown trout
3.3 饲料蛋白和脂肪水平对亚东鲑幼鱼肝脏生化指标的影响
在肝脏中, GOT和GPT是活性最高的两种转氨酶, 广泛存在于组织细胞线粒体内, 是一项反应肝功能的重要指标[34]。当肝细胞坏死或肝脏功能受到损伤时, 肝脏GOT和GPT活性下降, 血清和组织中的酶活性升高[35,36]。在本试验中, 在同一脂肪水平下, 随饲料蛋白水平增加, 肝脏GPT活性先升高后降低, 在46%蛋白水平时达到最大。在石英等[37]对血鹦鹉(Cichlasomasp.)的研究中, 随着饲料蛋白从37%升高到49%, 肝脏GPT也呈现先上升后下降的趋势。桑永明等[32]对方正银鲫的研究也有相似结果。在一定的饲料蛋白水平范围内, 肝脏GPT活性的增加, 意味着转氨作用的增强, 可以促进体内的氨基酸代谢。
甘油三酯和胆固醇含量是反映机体对脂肪代谢状况的重要指标, 二者均在肝脏进行合成[38]。在本试验中, 在同一脂肪水平下, 肝脏甘油三酯和胆固醇含量均随着饲料蛋白水平的升高呈现先上升后下降的趋势, 其原因可能因为过多的蛋白水平,增加了鱼体的代谢负担从而抑制了甘油三酯和胆固醇等脂肪代谢。在中高蛋白水平下(12%, 16%),饲料脂肪水平的升高, 降低了胆固醇含量, 其原因可能是饲料中过多的脂肪造成了脂肪肝, 影响了肝脏的正常功能。这在陈建明等[39]、Lin等[40]的研究中也有类似报道。
4 结论
在本实验条件下, 综合考虑饲料系数、增重率、成活率和消化酶活性等因素, 亚东鲑幼鱼饲料中粗蛋白和粗脂肪的适宜水平分别为46%和12%。
