人胚胎干细胞RNA的细胞质和细胞核内的定位
2020-06-09周凡琦冯文新谭普文马艳妮
周凡琦,冯文新,谭普文,马艳妮,王 栋,余 佳*
(1.中国医学科学院基础医学研究所 北京协和医学院基础学院 医学分子生物学国家重点实验室, 北京 100005;2.河南地矿职业学院, 河南 郑州 450000;3.南方医科大学基础学院 生物信息系, 广东 广州 510000)
人胚胎干细胞(human embryonic stem cells,hESCs)是研究早期胚胎发育的细胞模型。人受精卵在受精后5 d内形成胚泡期胚胎,hESCs从胚泡期胚胎的内细胞群中分离出来[1-2],hESCs具有无限的复制潜力和多能性,能够分化成人体内任何体细胞类型[1]。这一特性使其有机会更新非功能性损伤组织,如帕金森病、脊髓灰质炎等疾病,目前基于hESCs治疗多种疾病的研究在临床前阶段已经取得了十分不错的成绩[3-4]。
RNA定位在细胞发育、命运决定等方面具有重要作用,如酵母细胞在分裂时,ASH1 mRNA定位于子细胞芽尖,进行局部翻译以调控其交配型的选择,从而确保子细胞产生与母细胞不同的交配类型[5-6]。在果蝇胚胎中,Bicoid mRNA定位于胚胎前极,Oskar mRNA、Nanos mRNA定位在后极,这种定位模式有助于果蝇胚胎建立形态梯度,为发育中的胚胎形成合适的空间模式奠定基础[7-9]等。这些mRNA及其蛋白产物在特定的亚细胞位置中发挥独特的功能,对于细胞正常的生理活动具有极为重要的意义,同时越来越多的研究表明,mRNA特定亚细胞的定位现象十分普遍。对果蝇3 000多个转录本进行高分辨率免疫荧光分析,结果显示约有71%的mRNA在早期胚胎中表现出显著的亚细胞定位模式[10]。
目前RNA定位的研究主要集中研究果蝇胚胎或酵母细胞中RNA广谱定位,揭示低等生命及哺乳动物细胞中某些特定RNA的定位及机制。哺乳动物细胞中对广谱RNA定位及其机制的研究较少,暂无研究报道hESCs中RNA细胞核和细胞质定位图谱。本研究主要探讨hESCs细胞核和细胞质中RNA定位详情及其特点。
1 材料与方法
1.1 材料
1.1.1 细胞:hESCs为H1细胞系(中国医学科学院血液学研究所赠送)。
1.1.2 主要试剂:matrigel(Corning公司);DPBS(Gibco公司);mTeSR1、ReLeSR和Accutase(STEMCELL公司);核质分离试剂盒(Thermo Fisher Scientificals公司:NE-PER Nuclear and Cytoplasmic Extraction Reagents);Trizol、M-MLV反转录试剂盒(Invitrogen公司);SYBR GREEN mix(全式金生物科技有限公司);抗HSP90抗体、抗Fibrillin抗体(Proteintech公司);山羊抗兔、山羊抗小鼠二抗(上海碧云天生物有限公司);引物(天一辉远测序公司);建库及测序(诺禾致源公司)。
1.2 方法
1.2.1 hESCs体外培养:将matrigel铺至6孔板4 ℃过夜后接种H1克隆,使用mTeSR1培养。观察密度,4~6 d传代1次:DPBS清洗细胞,加入1 mL ReLeSR室温静置1 min后吸净,将细胞放至37 ℃孵箱中6~8 min,加入3 mL mTeSR1将克隆重悬,传代至6孔板中进行培养。在培养过程中,DPBS、mTeSR1均需37 ℃预热并且每天换液。
核质的分离:使用Accutase将H1克隆消化成单细胞,吸取10% input后1 000 r/min离心得到细胞沉淀。使用核质分离试剂盒,按照细胞体积加入适量预冷CER1重悬细胞,涡旋振荡15 s,冰上孵育10 min;加入预冷CER2,涡旋振荡5 s,冰上孵育1 min;涡旋振荡5 s后21 000×g离心5 min,立即转移上清至干净的EP离心管中,此为细胞质成分(cytoplasmic extraction,CE);向剩余沉淀部分中加入适量预冷NER,涡旋振荡15 s后冰上孵育40 min,且每10 min振荡混匀15 s;21 000×g离心10 min,转移上清至干净的EP离心管管中,此为细胞核成分(soluble nuclear extraction,SNE),SNE成分中不包含染色质部分。
1.2.2 Western blot检测细胞核与细胞质标志蛋白的表达:提取input、CE、SNE蛋白后进行SDS-PAGE电泳,湿转法转到PVDF膜上,5%脱脂牛奶室温封闭1 h,一抗4 ℃孵育过夜:HSP90、Fibrillin。TBST洗涤后加入二抗室温孵育40 min,TBST洗膜后,用化学发光法显影。
1.2.3 RT-qPCR检测细胞核与细胞质标志基因的表达:使用Trizol法提取RNA,采用M-MLV及Random primer法进行反转录,得到cDNA。RT-qPCR反应体系为:SYBR GREEN mix 5 μL,上下游引物(10 μmol/L)0.2 μL,cDNA 0.5 μL,去离子水4.1 μL;反应条件为:95 ℃预变性5 min;95 ℃变性20 s,60 ℃退火20 s,72 ℃延伸30 s,共40个循环;溶解曲线生成的反应程序为:95 ℃ 15 s,60 ℃ 15 s,95 ℃ 15 s。引物序列(表1)。
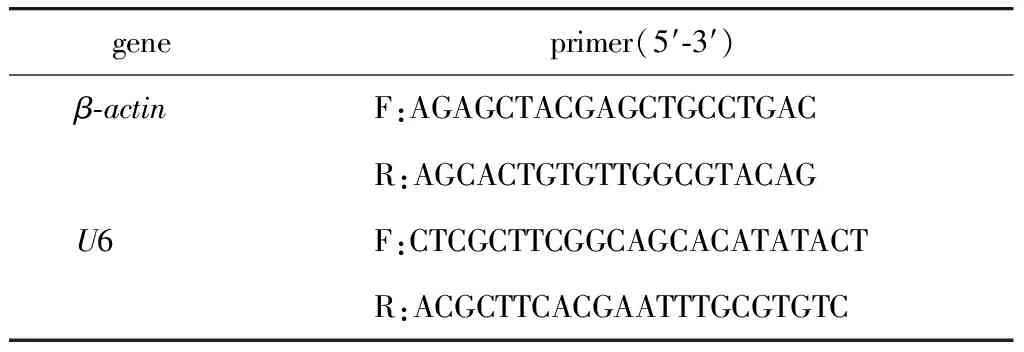
表1 RT-qPCR引物Table 1 Primer sequences for RT-qPCR
1.2.4 建库测序及生物信息学分析:将1 μg input/CE/SNE 总RNA去除rRNA并纯化回收,而后进行链特异性建库及测序;每组样品均有两个重复组;建库及测序项目编号为P101SC18060916-01。
使用Trimmomatics(v0.36)对原始测序数据进行质量控制,利用Hisat2(v2.0.5)将原始数据比对至GENCODE(v28)参考基因组(mm10)后,利用featurecounts计算基因表达量并计算TPM值。进一步利用edgeR计算差异表达基因以及挖掘亚细胞组分特异基因,Metascape被用于功能富集分析(gene ontology,GO),并使用pheatmap包绘制热图。
1.3 统计学分析
2 结果
2.1 hESCs核质的分离
分离hESCs细胞核与细胞质后检测其标志基因可见细胞质标志基因HSP90蛋白与β-actin RNA主要定位在细胞质中(P<0.001);细胞核标志基因Fribillin蛋白与U6主要定位于细胞核中(P<0.001)(图1)。
2.2 hESCs中RNA核质定位图谱
hESCs核质中总RNA的测序结果显示,细胞质中共定位30 232个RNA(TPM>1),这些RNA类型丰富,共57种RNA,其中34%为蛋白编码基因,17%为加工假基因,12%为基因间长链非编码RNA(long intergenic non-coding RNA,lincRNA)(图2A)。细胞核中共定位34 390个RNA(TPM>1),共31种RNA,其中74%为蛋白编码基因,7%为lincRNA,6%为反义RNA(图2B)。
尽管很多RNA同时定位于胞质与核质,但也存在特异性定位于胞质或核质的RNA。hESCs细胞质中特异定位的RNA多于细胞核(图3C),其中细胞质中特异性定位2 952个RNA,约91.8%为mRNA,5.22%为加工假基因(图3A);细胞核中特异性定位RNA共634个,其中41.42%为mRNA,21.89%为lincRNA(图3B)。这些基因的GO结果显示(图3D),细胞质中RNA功能丰富,参与细胞多种生理功能,如参与合成核糖核蛋白复合物、核苷酸代谢、氨基酸代谢、蛋白质折叠、细胞分裂等,在细胞的正常生理活动中起重要作用。细胞核中特异定位的RNA较少,其富集的GO条目也较少,包含钾离子反应、非钠离子依赖的有机阴离子运输、减数分裂重组等核内生理活动相关调控。可见hESCs细胞核和细胞质中特异性定位的RNA功能特点与其定位特点紧密相关。
2.3 RT-qPCR验证
根据上述分析结果各选择10个细胞核和细胞质特异定位的RNA进行验证,验证结果与测序结果一致(P<0.05)(图4),表明测序及分析结果真实可信(图4A)。细胞质中不仅特异性定位mRNA,如EEF2、HMGA1、LIN28A;此外还有一些非编码RNA(non-coding RNA,ncRNA)同样定位于此,如EEF1A1P5、TUBBP1、RPS28P7等(图4B)。除ncRNA外,细胞核中也有mRNA的定位,如自然杀伤细胞激活性受体(natural killer cell triggering receptor,NKTR)。
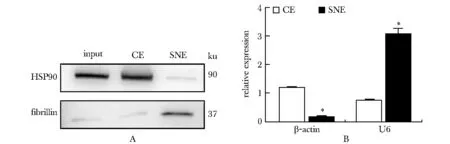
A.Western blot assay for markers(HSP90 and Fibrillin); B.RT-qPCR detection of marker genes(β-actin and U6);*P<0.001 compared with CE
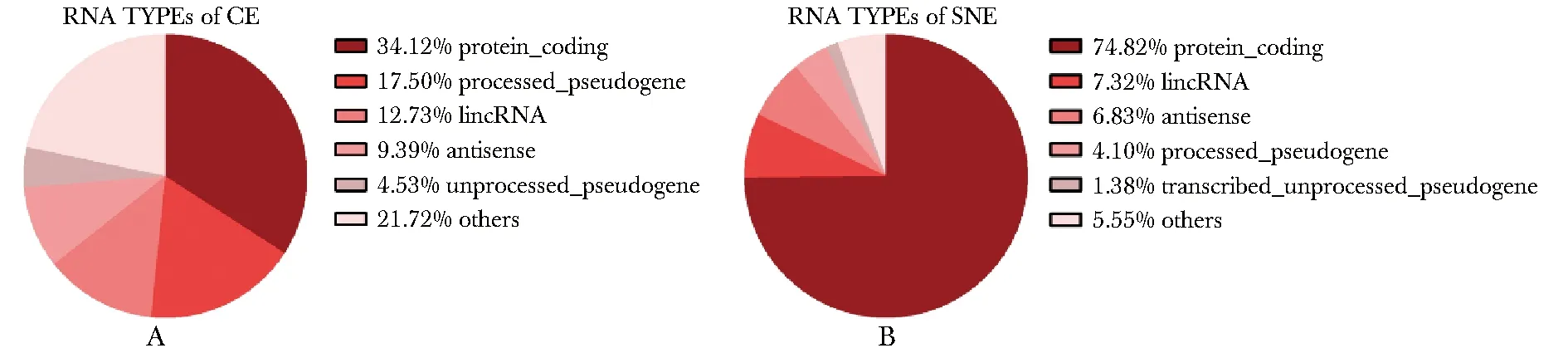
A.TOP5 of RNA types in CE; B.TOP5 of RNA types in SNE图2 hESCs细胞核和细胞质中RNA类型Fig 2 RNA types of cytoplasm and nucleus of hESCs
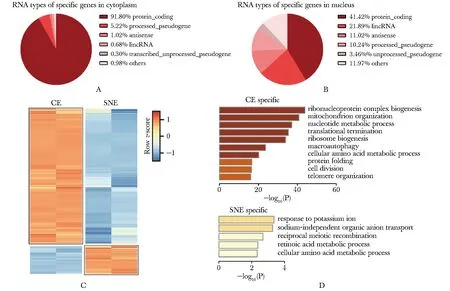
A.TOP5 of RNA types of specifically localized in cytoplasm; B.TOP5 of RNA types of specifically localized in nucleus; C.heatmap of specific transcripts between subcellular fractions; Colors indicate the percentile of the relative expression level; D.gene ontology(GO) enrichment analysis of specific transcripts between subcellular fractions; biological duplicates
图3 hESCs中RNA细胞核和细胞质定位图谱
Fig 3 The localization of RNA in cytoplasm and nucleus of hESCs(n=2)
3 讨论
研究者既往工作表明在胚胎发育过程中,RNA的精准定位及其定位的变化均具有重要调控作用。如在果蝇发育的胚胎期,orb mRNA逐渐在胚胎后极的细胞核周围形成独特的环状结构;同时,grp mRNA由弥散状态逐渐定位至胚胎后极以发挥其功能[8,11]。hESCs具有分化为人体内220多种细胞类型的潜力,它的分离与体外培养具有极其重要的基础研究和临床应用价值,明确hESCs的RNA定位图谱对于探索hESCs全能性的维持及研究RNA定位的生理功能具有重要意义。

A.RT-qPCR detection of the expression of cytoplasmic localized RNAs,**P<0.001 compared with SNE; B.RT-qPCR detection of the expression of soluble nuclear localized RNAs,*P<0.05,**P<0.001 compared with CE
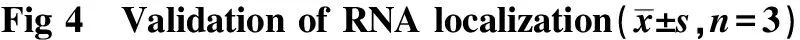
目前研究表明, RNA定位并局部翻译的调控模式在细胞生命活动中具有极大优势,首先,使基因表达在细胞质中受空间限制;其次,在局部刺激下,mRNA精准定位可在刺激原位迅速调控翻译水平,高效实现细胞对刺激的应答反应;第三,RNA定位的模式对于细胞来说十分经济,定位于亚细胞的mRNA可被多次翻译,产生大量蛋白质[12]。由此可见,RNA精准定位对于细胞正常生理功能具有重要意义。
本研究初步描绘了hESCs中RNA的细胞核和细胞质定位图谱,揭示了hESCs中6万多个RNA的核质定位详情及特点,如细胞质中RNA类型丰富,可能由于细胞质为RNA翻译、RNA与蛋白质发挥功能的主要区域,大多数mRNA转录后被运输至细胞质进行翻译;也有很多ncRNA在胞质中可能发挥调控功能。而细胞核中定位大量mRNA,分析原因可能是转录后正在进行加工以及转运出核的RNA;也有一些成熟的mRNA定位于胞核中,这些mRNA可能是作为“备用池”(reserve pools)发挥功能,在细胞中有一些基因的转录以爆发的形式发生,而将这部分mRNA储存于核中可以缓冲胞质中这些转录本的水平,从而减少转录噪声带来的基因表达变异[13];此外猜测可能有一部分mRNA在细胞核中具有某些调控功能。这些RNA定位特点对既往RNA核质定位认知进行了一定的补充。
同时发现特异性定位于细胞质的RNA多于细胞核,且这些RNA类型与整体定位的RNA类型不同,特异性定位于胞质中的mRNA占比很高,超过90%;而细胞核中也有很多mRNA特异性定位于此,占核中特异性RNA的41%,这些mRNA翻译的蛋白可能更倾向于定位至核散斑与核浆处来发挥功能[14],也可能存在未知的调控功能,如NKTR在自然杀伤细胞(natural killer cell,NK)中编码膜锚定的蛋白,存在于NK细胞表面促进它们与靶细胞的结合,但NKTR在hESCs中定位于细胞核,这提示该mRNA可能在hESCs中具有未知的功能。可见mRNA、ncRNA的核质定位不由RNA类型决定,更可能是通过其自身序列特点、结构特点或其他调控因子等调控其定位。
目前研究表明RNA的核质定位可通过多种因素共同调控实现,如RNA序列中的定位元件可调控RNA出核或滞留核中,如大部分mRNA的3′UTR区有出核序列,而很多长链非编码RNA中具有核滞留顺式元件,这些序列可被特定的RNA结合蛋白(RNA binding protein,RBP)识别并结合,调控RNA出核或核滞留,使其准确定位并发挥功能。因此RNA的定位元件与转运RBP将共同调控RNA的定位,此外RNA的一些修饰水平、RNA选择性剪接、RNA3′端选择性加尾等也将影响RNA的核质定位[15]。研究这些RNA特异定位在细胞核与细胞质的机制是本课题进一步的研究方向。
综上,本研究展示了很多有趣新颖的RNA在细胞核和细胞质定位,全面描绘了hESCs细胞核和细胞质中RNA定位图谱,为进一步探索hESCs中RNA精确的定位图谱提供了基础。
