干旱-高钙对麻栎幼苗非结构性碳水化合物含量和分配的影响
2020-06-03李亚楠张淞著张藤子魏亚伟周永斌朱文旭
李亚楠,张淞著,2,张藤子,魏亚伟,2,李 慧,2,周永斌,2,朱文旭,2,殷 有,2,*
1 沈阳农业大学林学院, 沈阳 110866 2 辽河平原森林生态站, 昌图 112518
非结构性碳水化合物(Non-structural carbohydrate,NSC)是植物生长代谢过程中重要的能量供应物质,其在植物体中含量的变化在很大程度上影响着植株的生长状况及应对逆境的能力[1- 2],近年来碳饥饿假说更是成为植物干旱生理死亡过程的研究热点[3]。但干旱环境中树木NSC变化的研究结果并不一致[1- 2,4- 6],有研究认为干旱过程中树木关闭气孔以阻止细胞快速失水,但同时抑制光合作用,碳同化量少于碳呼吸量,造成树木碳的负平衡[1- 2,7- 8]。也有研究表明树木可以在干旱过程中通过降低生长等自身调节策略保持碳平衡[6,9-10]。
目前干旱-半干旱区树木碳平衡的研究只关注干旱对树木NSC的影响,对于干旱-半干旱区特有的环境因子与干旱共同作用下,树木NSC变化趋势的研究较少[11]。事实上在半干旱区影响植物生长的环境因子中,除水分被认为是限制人工林生长的主要限制因子外[12- 15],广泛分布的、以高钙为主要特征的土壤钙积层,成为限制树木生长的另一重要环境因子[15-17]。钙作为重要的第二信使,已被证明是能够调节气孔关闭的关键信号物质[18-19]。高钙能够调节树木气孔关闭从而影响光合作用,并最终影响水分利用效率和生长速率[20- 21]。鉴于高钙土壤分布的广泛性[22- 23]和钙素在调控植物气孔关闭过程中的关键作用[24- 25],阐明高钙与干旱共同作用对树木NSC的含量和分配的影响,推测干旱和高钙环境下树木发生碳饥饿的风险,对于理解树木在干旱半干旱区特殊环境下的生理过程具有重要意义。
麻栎(Quercusacutissima)属壳斗科栎属的重要人工林树种,耐干旱瘠薄,能稳定表土、缓冲径流、拦截泥沙、是营造水土保持林的优良树种[26- 27],本研究以麻栎幼苗为研究对象,研究干旱和干旱-高钙环境中麻栎生长、光合、NSC含量及其在不同器官中分配的变化,探究干旱-高钙环境下麻栎发生碳饥饿的可能性。
1 材料与方法
1.1 试验材料与设计
试验在沈阳农业大学林学院北山科研基地进行。试验前将麻栎种子用5%次氯酸钠消毒15 min,无菌水冲洗3—5次,于25℃培养箱中培养,露白后挑选发芽一致的种子进行播种。试验所用塑料盆规格为底径14 cm,高16 cm,每盆装灭菌土1.5 kg,供试土壤采集自沈阳农业大学后山杂木林,土壤碱解氮、速效磷、速效钾含量分别为339.50、12.07、183.40 mg/kg,土壤水溶性钙和交换性钙含量为235.65、4579.95 mg/kg,土壤最大持水量是31.70%。
试验设置3个处理,分别为对照组(Control,CK),即正常供水(最大持水量的80%±5%),不加钙;干旱处理组(Drought,DC),即供水量减至最大持水量的40%±5%,不加钙;干旱-高钙处理组(Drought and high calcium,DH),即供水量为土壤最大持水量的40%±5%并添加800 mg/kg的Ca2+,每个处理设6个重复。根据预实验,高钙处理添加的钙以氯化钙水溶液的形式添加到土壤。
2017年5月12日挑选发芽一致的种子进行播种,每盆播种3粒种子,待种子出土1周后进行间苗,每盆保留一株长势一致的幼苗。间苗前进行合理的水分管理,以确保幼苗的正常生长。间苗后进行钙处理并在土壤含水量降至目标水量后开始水分控制,用称重法每两天测定花盆重量,记录不同处理耗水量并根据土壤耗水量添加水分,整个试验过程中所添加的水分均为去离子水。在水分处理3个月后结束试验,进行相关指标测定。
1.2 指标测定
1.2.1光合参数的测定
选择2017年6月10日(即干旱处理一周后)和9月11日晴朗的上午,使用Li-COR公司的Li- 6400便携式光合测定仪测定麻栎幼苗树梢第三片完全展开的健康叶片,叶室配有LED红蓝光源,有效光辐射强度(Photosynthetically active radiation,PAR)设置在1000 μmol m-2s-1,每个处理随机选择3个重复。测定的光合参数为净光合速率(Net photosynthetic rate,Pn,μmol m-2s-1)和气孔导度(Stomatal conductance,Gs,mol m-2s-1)。
1.2.2植物样品的采集
在生长期末尾落叶前进行植物样品的采集,将收获的植株清洗干净吸干植株表面水后分为根、茎、叶放入信封,烘箱105℃杀青30 min,随后65℃烘至恒重,使用万分之一天平测定植株根、茎、叶生物量。
1.2.3NSC的测定
烘干后的植物各部分样品粉碎后,过100目筛,装入自封袋标记备用。称取干样50 mg,放入10 mL离心管,加入4 mL 80%乙醇,80℃水浴30 min,3000 r/min离心10 min,将上清液倒入刻度试管中,其残渣加入2 mL 80%乙醇重复提取2次,合并上清液,上清液经活性炭80℃脱色,定容后即为可溶性糖待测液,蒽酮比色法测定625 nm波长下的吸光度,根据标准曲线求出提取液中糖的含量;在提取可溶性糖后的沉淀加蒸馏水糊化15 min,随后用9.2 mol/L高氯酸2 mL提取15 min后离心,收集上清液,再向沉淀中加入4.6 mol/L高氯酸2 mL提取15 min后离心,合并上清液,用蒽酮比色法测定625 nm波长下的吸光度,根据标准曲线求出提取液中糖的含量,在计算淀粉含量时将所得糖含量乘以0.9作为扣除水量后的实际淀粉含量。非结构性碳水化合物以可溶性糖和淀粉的总和表示。
1.3 数据处理
植株平均可溶性糖或淀粉或非结构性碳水化合物含量(mg/g)=(根可溶性糖或淀粉或非结构性碳水化合物含量×根系生物量+茎可溶性糖或淀粉或非结构性碳水化合物含量×茎生物量+叶片可溶性糖或淀粉或非结构性碳水化合物含量×叶片生物量)/整株生物量。用SPSS 19.0和Excel 2016进行数据统计分析,采用单因素ANOVA分析不同处理间光合参数和非结构性碳水化物含量的显著性差异(P<0.05)。
2 结果与分析
2.1 不同处理对麻栎幼苗生长及光合参数的影响
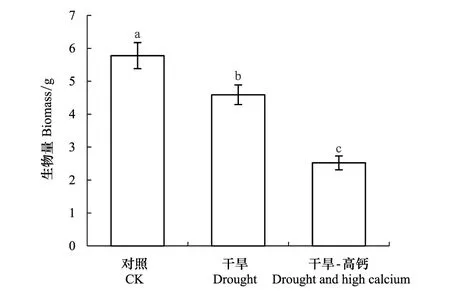
图1 麻栎幼苗生物量Fig.1 Biomass of Q. acutissima seedlings 对照CK:Control,即正常供水不添加钙;干旱DC:Drought,即供水量减至最大持水量的40%±5%,不加钙;干旱-高钙DH:Drought and high calcium,即供水量为土壤最大持水量的40%±5%并添加800 mg kg-1 Ca2+;不同小写字母表示处理麻栎幼苗间生物量差异显著(P<0.05)
图1表明,干旱和干旱-高钙处理下麻栎幼苗的生长都受到抑制,而干旱-高钙对麻栎幼苗生长的限制更显著(P<0.05)。干旱胁迫下麻栎幼苗生物量较对照显著降低15.95%(P<0.05)。干旱-高钙处理下麻栎幼苗生物量较对照和干旱处理均显著降低,分别下降56.38%和48.10%。
从图2可见,干旱初期,干旱及干旱-高钙处理,麻栎幼苗叶片净光合速率较对照分别下降17.07%和49.44%,其中干旱-高钙处理下叶片净光合速率和气孔导度下降显著;气孔导度较对照分别下降40.14%和58.42%,显著降低。与单一干旱相比,干旱初期干旱-高钙处理显著降低了麻栎幼苗叶片净光合速率和气孔导度,分别下降39.03%和30.54%。干旱持续3个月后,干旱及干旱-高钙处理下麻栎叶片净光合速率和气孔导度显著降低,其中净光合速率分别下降93.36%和93.31%,气孔导度分别下降58.33%和63.78%。而干旱和干旱-高钙处理间叶片净光合速率和气孔导度差异不大。
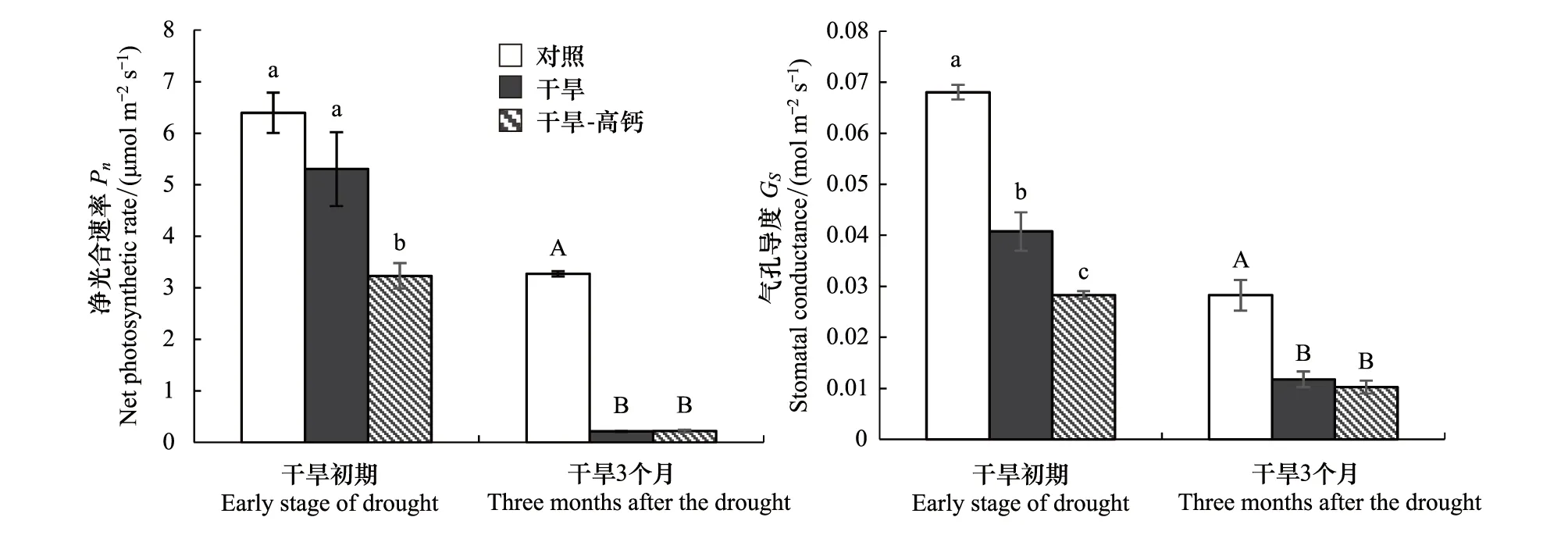
图2 麻栎幼苗光合参数Fig.2 Photosynthetic characteristics of Q. acutissima seedlings不同小写字母表示干旱初期处理间差异显著,不同大写字母表示干旱3个月处理间差异显著(P<0.05)
2.2 不同处理对麻栎幼苗NSC含量和分配的影响
从表1可见,麻栎幼苗的平均可溶性糖含量在干旱和干旱-高钙处理下较对照分别增加了15.38%(P<0.05)和15.56%(P<0.05),但是两处理不同器官可溶性糖含量却有不同的变化趋势:与对照相比,干旱处理的麻栎幼苗茎和叶可溶性糖含量均增加,仅根系可溶性糖含量降低,但干旱对麻栎各个器官可溶性糖含量影响并不显著。干旱-高钙处理的麻栎幼苗根系及茎可溶性糖含量均降低,其中根系可溶性糖含量显著下降,仅叶片可溶性糖含量显著增加。与单一干旱处理相比,干旱-高钙处理显著降低了根系及茎可溶性糖含量,分别下降53.18%和44.44%。叶片及植株平均可溶性糖含量分别增加19.66%和0.17%,增加不明显。
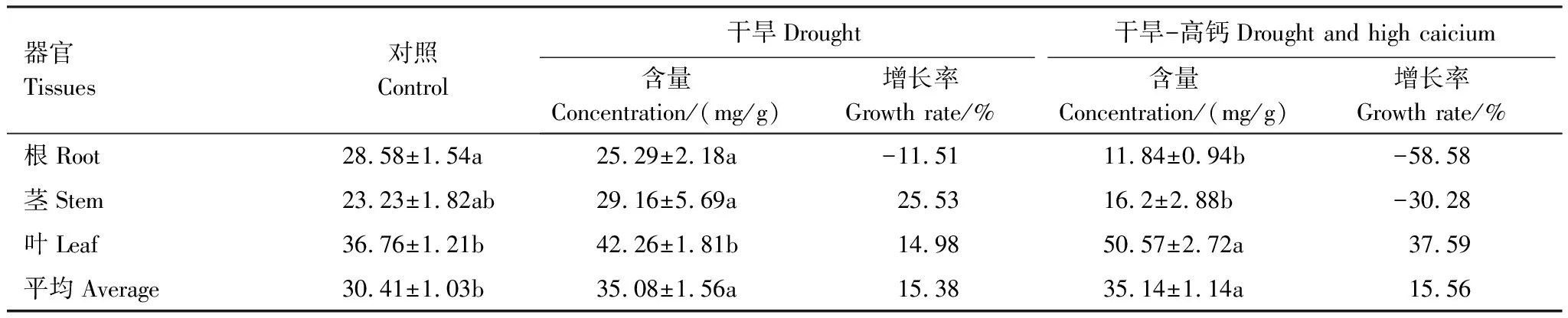
表1 麻栎幼苗各个器官可溶性糖含量
不同小写字母表示不同处理间麻栎幼苗各个器官可溶性糖含量差异显著(P<0.05)
表2表明,麻栎幼苗根系、茎及植株平均淀粉含量在干旱处理时达到最高,干旱-高钙处理下淀粉含量最低,叶片淀粉含量随对照、干旱及干旱-高钙处理依次增加。与对照相比,干旱处理下麻栎幼苗根系、茎、叶及植株平均淀粉含量显著增加;干旱-高钙处理降低了根系及植株平均淀粉含量,叶片淀粉含量显著增加。与单一干旱相比,干旱-高钙处理下麻栎幼苗根系、茎及植株平均淀粉含量分别下降57.71%、61.16%和55.84%,显著降低;叶片淀粉含量增加2.84%,变化不显著。

表2 麻栎幼苗各个器官淀粉含量
不同小写字母表示不同处理间麻栎幼苗各个器官淀粉含量差异显著(P<0.05)
由表3可知,与对照相比,干旱处理麻栎幼苗根、茎、叶片及植株平均NSC含量均显著增加,干旱-高钙处理下麻栎幼苗根系、茎及植株平均NSC含量显著降低,叶片NSC含量显著增加。干旱-高钙处理下,麻栎幼苗根系、茎及植株平均NSC含量分别下降57.05%、57.54%和41.11%,显著降低;叶片NSC含量显著增加14.52%。

表3 麻栎幼苗各个器官NSC含量
不同小写字母表示不同处理间麻栎幼苗各个器官NSC含量差异显著(P<0.05)
3 讨论
3.1 NSC含量
干旱处理的麻栎幼苗平均NSC含量较对照显著升高,出现NSC积累的现象。树木在干旱条件下碳同化量下降,但同时生长速率的降低减少了对碳的需求,当树木生长速率下降的速率快于光合速率降低的速率时,光合产物更多的以NSC的形式储存[4,28]。一些研究者认为NSC积累是被动过程,尤其是淀粉的积累对植物抗旱性并无帮助[3],但是逆境条件下植物提高NSC储存量是植物应对逆境的生态对策之一,同时组织中NSC含量的变化在很大程度上影响着树木应对逆境的能力以及逆境后恢复生长的能力[2],因此NSC积累是树木适应干旱的有效生态对策之一。
干旱和高钙共同作用下麻栎幼苗平均NSC含量较对照和干旱处理均显著降低。干旱-高钙处理降低NSC含量的原因之一在于能够更显著地降低光合作用(图2)。干旱会导致气孔关闭,降低光合底物二氧化碳的吸收,同时损伤叶肉细胞,降低光合酶活性,从而降低光合速率[29- 31]。钙元素是调控气孔关闭的信号物质,高钙能够诱导气孔关闭,从而降低光合作用[19- 21]。本研究中干旱和干旱-高钙处理3个月后均显著降低了麻栎光合作用,但是在实验处理初期,干旱-高钙处理较对照已经显著地降低麻栎光合作用(图2),造成光合产物供给大量减少,从而在显著抑制麻栎生长的同时降低NSC含量。另外,钙离子是植物重要的第二信使,调控众多生理过程[32- 33],NSC含量的降低可能与过量钙离子调控的生理过程有关,但是对于过多的钙离子如何影响能量(NSC)储存、呼吸作用和生长等影响NSC含量的生理过程,目前尚不明确。
3.2 NSC分配
干旱处理的麻栎幼苗中,除根部可溶性糖略有减少外,麻栎幼苗各器官可溶性糖、淀粉、NSC含量都增加,这与雷虹等[28]研究结果一致,可溶性糖是光合产物的运输媒介[34],可作为生长的原料或转化为淀粉储存[3],而干旱条件下树木生长受限可溶性糖得到积累并部分转化为淀粉[35]。根系可溶性糖降低原因可能是干旱条件下生长量主要集中在根部[36],从而消耗了根系中的可溶性糖。
干旱-高钙处理改变了麻栎幼苗的NSC分配,叶片NSC含量显著升高,根和茎中的NSC含量却显著降低(表1—表3)。表明干旱-高钙处理抑制了NSC从碳源(叶片)到碳汇(根)的运输,导致碳源组织中NSC积累而碳汇组织NSC降低出现碳负平衡。张海平[37]在研究花生时也发现高钙条件下叶中会出现淀粉累积。但是具体生理机制尚缺少系统阐明,在后续研究中需进一步研究。NSC对于植物适应逆境以及逆境后的恢复具有重要意义,对于落叶树种,叶中的NSC将随着落叶损失掉,根和茎中的NSC才是其度过休眠期后再次萌发生长的主要能量来源,根和茎中NSC的减少将抑制植物适应逆境的能力。
3.3 干旱-高钙导致植物碳饥饿的可能性
干旱条件下,树木NSC变化趋势随树种不同而差异很大[38]。碳饥饿假说认为在长期干旱条件下,树木呼吸作用大于光合作用,出现NSC含量减少,最终使得NSC含量无法支持树木代谢而死亡。而极端干旱条件下,树木往往在未消耗掉储存的NSC之前就由于水力失败而死亡,因此树木的碳饥饿往往是在长期干旱过程中发生的[39- 41]。因此,在更长时间尺度下比较干旱对麻栎幼苗NSC变化动态的影响能够更好的反应麻栎幼苗在干旱条件下出现碳饥饿的可能性,即使本研究中单一干旱条件下麻栎幼苗生长降低,NSC含量升高的现象,随着干旱持续时间的增加,碳的负平衡同样会出现,存在碳饥饿的风险。然而干旱-高钙环境下,麻栎幼苗平均NSC含量较对照显著减少,表明干旱-高钙处理缩短了树木保持碳平衡的时间,而干旱和高钙是干旱半干旱地区树木自然生长过程中需要长期客观面对的环境胁迫,因此树木在高钙土壤上发生碳饥饿的风险更高。
另外Sala等人认为碳饥饿的过程可能由于非结构性碳水化合物的传输受阻,而导致树木局部器官发生碳饥饿[42]。干旱-高钙处理的麻栎幼苗NSC更多积累在叶中,根和茎中分配的NSC显著减少,如出现干旱落叶或树木进入休眠状态落叶,则会加剧树木NSC减少的趋势,增加树木发生碳饥饿致死的可能性。
4 结论
干旱-高钙处理减少了麻栎幼苗平均NSC含量的同时,抑制了NSC由叶向茎和根的运输,干旱-半干旱区高钙土壤上生长的树木更易发生碳饥饿死亡。
