甘氨酸脱羧酶基因增强子区(-4.2~-2.2 kb)基因图谱绘制及其活性检测
2020-06-03赵爽郑华川
赵爽,郑华川
(中国医科大学附属盛京医院实验肿瘤学实验室,辽宁 本溪 117004)
甘氨酸脱羧酶(GDC,P-protein)构成甘氨酸裂解系统,该系统还包含H蛋白,T蛋白和L-蛋白,上述蛋白的构成比例为2P:27H:9T:1L[1-2]。该系统催化脊椎动物体内甘氨酸裂解的初始阶段,产生CO2,NH3,N5,N10-亚甲基和NADH+H+,是甘氨酸代谢的主要途径;在植物研究中发现,叶线粒体含有大量的GDC蛋白,主要参与植物光合作用[2-3]593-597。GDC基因突变可导致人类血清甘氨酸累积,最终引起神经系统疾病发生,如非酮症性高甘氨酸血症(NKH);同时还参与肺癌的形成[4-5]。目前研究发现,所有的脊椎动物所有组织中,鸡与人类的GDC基因结构和功能相似度较高,且鸡体内甘氨酸裂解活性较强[6]。因此,我们克隆出了鸡GDC基因启动子前(-4.2~-2.2 kb)区域,绘制其基因图谱,检测其对GDC启动子的调节能力。
1 材料和方法
1.1 上游GDC基因KX DNA片段测绘
GDC基因5’端7.3 kb区域克隆自噬菌体λCPG301,再从GDC基因5’端7.3 kb区域分离DNA片段-4.2~-2.2 kb。将分离出的DNA片段插入KpnI和XbaI位点之间,并筛选出阳性克隆命名为PBS-KXb。利用KpnI-Pst1、KpnI-HincII、KpnI-HindIII、KpnI-EcoRI、KpnI-SacI、KpnI-XbaI、XbaI-PstI、XbaI-HindIII双酶切重组质粒PBS-KXb并进行琼脂糖电泳,拍照。
1.2 质粒构建
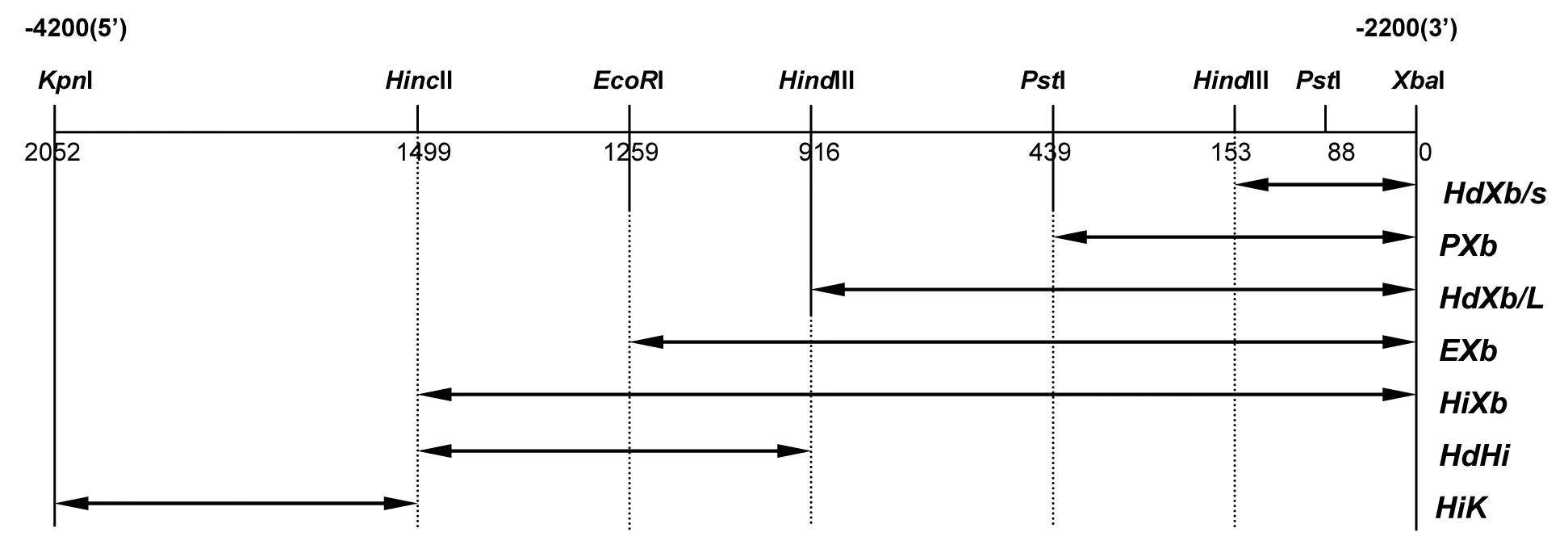
以pGLΔXbaI-KX(KpnI-XbaI)为载体构建KX区域的截断体。用HindⅢ-XbaⅠ、PstI-XbaI、EcoRI-XbaI、HincⅡ-XbaI、HincII-HindIII或HincⅡ-KpnI酶切KXB区域全长或部分DNA片段插入到pBS(-)质粒,并转化到大肠杆菌通过菌落PCR或限制性内切酶酶切筛选阳性克隆,利用CUGA测序试剂盒测序(日本genetech),测序引物:T3、T7、MB FW、XK-R-2、XK-R-3、XK-F-2。进一步构建pGLΔXbaI-KpnI-XhoI(KX)、HindⅢ-XhoⅠ(HDX/S)、PstI-XhoⅠ(PX)、HindⅢ-XhoⅠ(HDX/L)、EcoRI-XhoI(EX2)、HincⅡ-XhoI(HIX)、HindII-HincIII(HDHI/L)或KpnI-HindII(HIK)并大规模制备这些质粒DNA。
1.3 荧光素酶报告基因检测
肝癌细胞HepG2利用DMEM培养基加入10%胎牛血清培养到80%。胰酶消化,离心收集细胞沉淀,计数,15 000/孔铺入96孔板,利用转染试剂Lipo3000(lipofectamine3000)转染野生型或重组质粒pGL3-basic及pRL-SV40,48 h后,裂解细胞,利用双荧光素酶报告基因试剂盒(Promega)检测荧光素酶活性。
2 结 果
2.1 利用测序引物对KXb全长测序,结果见图1,KXb全长2052 bp,含有限制性内切酶KpnI、HincII、EcoRI、HindIII、PstI、XbaI。在760~790区域含有GATA重复序列。
2.2 本研究利用KpnI和XbaI双酶切的方法从GDC基因5’端上游-4200~-2200区域分离出2.0 kb长的DNA片段KXb,并将该片段插入到pBlueScript SK(+/-)载体中,成功构建重组质粒PBS-KXb,通过限制性内切酶酶切重组质粒发现GDC基因上游区域KXb段含有以下酶切位点KpnI、HincII、EcoRI、HindIII、PstI、XbaI。利用限制性内切酶位点构建KXb片段的截短体,并将截短体插入到载体pBlueScript SK(-)载体中,成功构建以下重组质粒pBS(-)KpnI-XbaI(KXb)、HindⅢ-XbaⅠ(HdXb/S)、PstI-XbaⅠ(PXb)、HindⅢ-XbaⅠ(HdXb/L)、EcoRI-XbaI(EXb)、HincⅡ-XbaI(HiXb)、HindII-HincIII(HdHi/L)或KpnI-HindII(Hik),见图2。
2.3 构建以下截断体的重组质粒pGLΔXbaI、KX(KpnI-XbaI)、HdX/S(HindⅢ-XbaⅠ)、PX(PstI-XbaⅠ)、HdX/L(HindⅢ-XbaⅠ)、EX2(EcoRI-XbaI)、HIX(HincⅡ-XbaI)、HDHI(HindII-HincIII)或HIk(KpnI-HindII),见图3。
2.4 将重组质粒(截短体)转染到肝癌细胞中,观察各截短体的荧光素酶活性,发现KX全长和HDHI/L,HIK两种截短体荧光素酶活性明显高于空载体pGLΔXbaI、HDX/L截短体只有轻微升高,其他四种截短体HDX/S、PX、EX2、HIX荧光素酶活性则低于空载体pGLΔXbaI,见图4。
3 讨 论
甘氨酸脱羧酶广泛存在于真核生物体内,植物体内的甘氨酸脱羧酶主要参与光合作用,哺乳动物体内的甘氨酸脱羧酶则主要参与甘氨酸代谢[2-3]593-597,3692-3697。现有研究发现,甘氨酸脱羧酶基因突变导致非酮症性高甘氨酸血症,这种疾病中,甘氨酸脱羧酶基因突变多出现在该基因的5’端,5’端突变可能影响甘氨酸脱羧酶基因的转录,从而影响甘氨酸脱羧酶的mRNA表达水平[5]e69。鸡GDC基因包括25个外显子,共3572个碱基,处于202503和240569之间,位于Z染色体负链上(登录号:AC202790)[6]3323-3329。UpSp1、InvSp1和下游CP2基序是顺式作用原件,可以启动鸡GDC基因转录,而5’-端区域-4155/-83包括KX、UpR1S、UpR1A、UpR2S和UpR2A区域则可能会负向调节启动子活性[7]。所以本实验主要研究KX(-4200~-2200)区域,通过测序结果发现,该区域确实含有CTTCTT重复序列,该序列可以通过结合蛋白,启动GDC基因转录[8]。但该区域含有的其他序列的作用还不明确,所以我们进一步构建了多种截短体来探究其调节作用。
通过双荧光素酶报告基因检测,我们发现鸡GDC基因5’-端区域-4200/-2200基因片段中,KX全长,HDHI和HIK截短体的荧光素酶活性升高,但是HDX/S、PX、EX2、HIX截短体的荧光素酶活性均有一定程度的降低,KX全长含有完整的调节元件,可以启动GDC基因表达,HDHI和HIK截短体区域则含有5′-CTTCTT-3′基序也可以启动GDC基因表达,与Hong S等的研究结果一致,均可以增强GDC基因启动子活性[8]10523-10532。但是HDX/S、PX、EX2、HIX截短体的荧光素酶活性降低,则说明HDX/S区段可能抑制GDC基因启动子活性。鸡GDC基因与人的GDC基因具有一定的相似度,且具有一些相同的原件,可以为人GDC基因的研究提供一定的依据。

图1 GDC基因上游KXb区域基因图谱
A
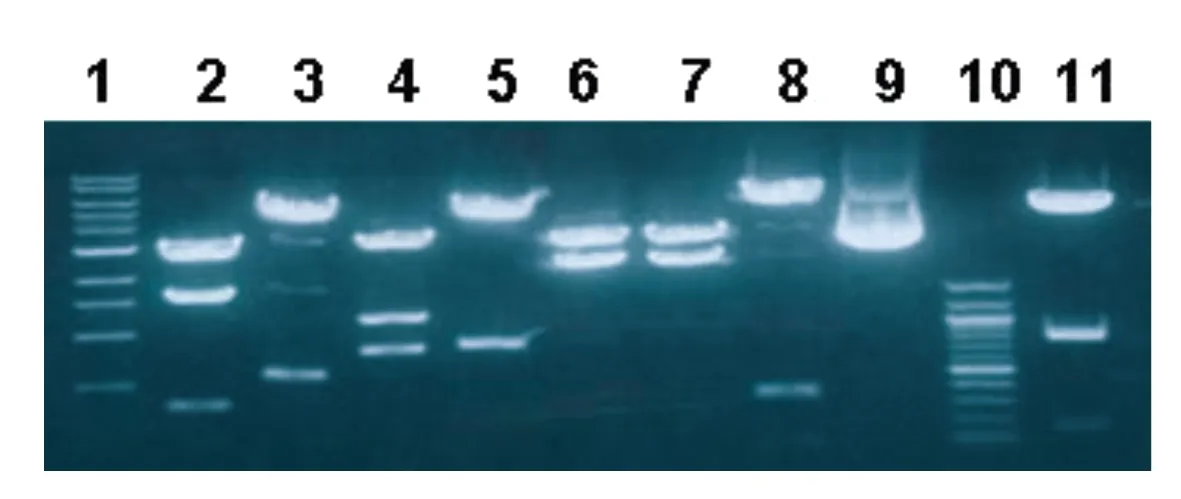
B
A:pBS-KX重组质粒双酶切琼脂糖电泳,1:1 kb maker;2:KpnI-Pst1;3:KpnI-HincII;4:KpnI-HindIII;5:KpnI-EcoRI;6:KpnI-SacI;7:KpnI-XbaI;8:XbaI-PstI;9:noncut;10:100 bp maker;11:XbaI-HindIII;B:GDC上游KXb区域酶切图谱
图2 GDC上游KXb区域酶切图谱

图3 GDC基因上游KXb区域截短体荧光素酶报告基因的构建
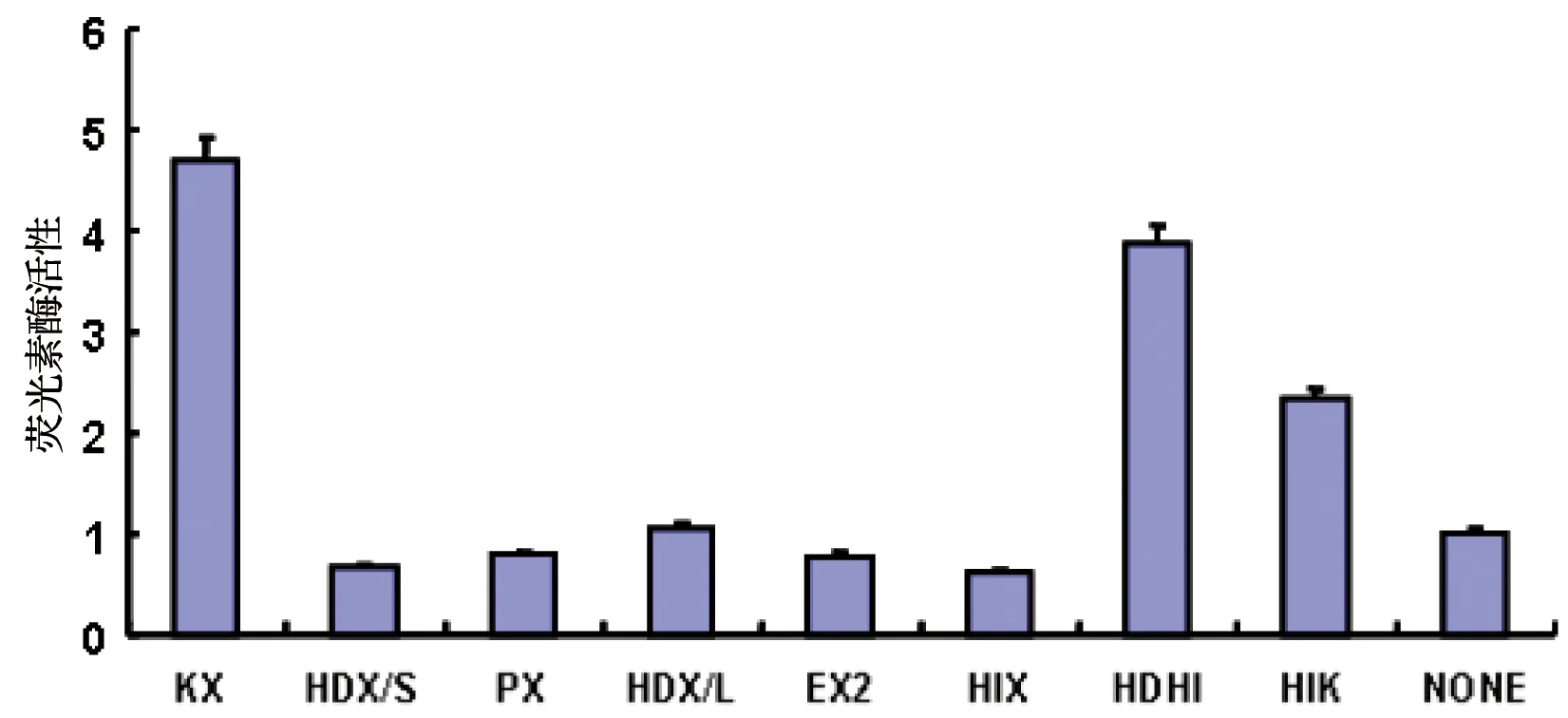
图4 GDC基因上游KXb区域截短体的启动活性检测
基因组DNA调取的方法有很多,long and accuracy PCR,genome walking及限制性内切酶法调取DNA[9],而本研究采用的是酶切法调取DNA,根据现有研究预测GDC基因的转录调控区域,最后我们利用软件查找转录调控区域内的酶切位点,选择KpnI和XbaI两种限制性内切酶,调取GDC基因5’-端区域-4200/-2200基因,并将该片段插入到PBS(-)载体中,通过单酶切检测片段长度,进一步用双酶切鉴定单酶切的结果,最后得出GDC基因KX区域的物理图谱。通过测序验证调取DNA的基因序列,绘制GDC基因KX区域的基因图谱。酶切法调取DNA的方法相较于其他方法,用时较短,不易发生基因突变,结果的可行度较高,但DNA片段必须为单一限制性内切酶,限制了该方法的使用。
