Soil-moisture dynamics and tree-water status in a Picea crassifolia forest,Qilian Mountains,China
2020-06-02HuLiuLinLiSiJiaWangQiYueYangWenZhiZhao
Hu Liu ,Lin Li ,3,SiJia Wang ,3,QiYue Yang ,WenZhi Zhao *
1.Linze Inland River Basin Research Station,Chinese Ecosystem Research Network,Lanzhou,Gansu 730000,China
2.Key Laboratory of Ecohydrology of Inland River Basin,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou,Gansu 730000,China
3.University of Chinese Academy of Sciences,Beijing 100029,China
ABSTRACT Landscapes of the mountainous regions in northwestern China comprise a unique pattern of vegetation,consisting of a mo‐saic of grassland and shrub-forest.Forests generally self-organize into ordered structures and coalesce into blocks on north-facing slopes or stripes along southeast-facing slopes,with Picea crassifolia being the most representative and domi‐nant tree species.We investigated the tree-water status and soil-moisture dynamics at a forest site(Guantan)of the Qilian Mountains in northwest China.The 30-minute-interval measurements of tree-sap flow during the growing season of 2008 are presented,and the potential functional relations between tree transpiration and environmental factors are evaluated.Soil moisture and solar energy were identified as the most influential factors,explaining more than 70%of the variance in sap flow.Based on field measurements obtained at the forest site,a stochastic model of soil-moisture dynamics was test‐ed;and the steady-state probability density functions(PDFs)of the long-term soil-moisture dynamics and static tree-water stress were estimated using the validated model and parameters.We found that the model reproduced measured soil mois‐ture well,despite all the simplifying assumptions.The generated PDF of long-term soil moisture was relatively open,with middle to low average values;and the calculated density of the static tree-water stress at the forest site was largely concen‐trated between 0 and 0.6,suggesting a moderate water-stress situation in most cases.We argue that both water and energy are limiting factors for vegetation at the forest site.In addition,the tradeoff between reduced evapotranspiration(ET)from limited solar energy and increased soil-moisture availability may create a stressed but tolerable environment and,in turn,produce a relatively constant ecological niche favorable to Picea crassifolia growth.
Keywords:sap flow;soil moisture;stochastic modeling;semiarid alpine ecosystem
1 Introduction
Arid inland river basins in Northwest China are unique ecosystems consisting of ice and snow,frozen soil,alpine vegetation,oases,deserts,and riparian for‐est landscapes,existing in a delicate ecohydrological balance(Liuet al.,2015).This region occupies nearly one-quarter of the country's total land area,and runoff from the mountains is its only water source(Mogha‐das,2008;Xuet al.,2009).Landscapes of the alpine ecosystems in this region usually present a unique pat‐tern of vegetation,consisting of grassland matrices and shrub-forest mosaics(e.g.,Tianshan Mountains,Qilian Mountains;Figures 1a,1b—a landscape tradi‐tionally thought to play a critical role in water-source conservation(Xuet al.,2009;Tianet al.,2011).For‐ests generally self-organize into ordered structures and coalesce into blocks on shady slopes(north-facing)or stripes along half-shady slopes(southeast-facing)in the altitude range of 2,600 to 3,200 m(Zhaoet al.,2007).Because these forests can play an important role in soil and water conservation for the headwater region of the inland river basins,the Chinese government allo‐cated large annual budgets for their protection,recov‐ery,and revegetation over the past decades(Kanget al.,2007;Andersen and Yang,2010).Despite the rec‐ognition of these forests'importance,however,relative‐ly little research has been done on even the most basic questions,such as what causes these vegetation pat‐terns to develop.Understanding the ecohydrological mechanisms for vegetation patterns(i.e.,relationships between the plants and water)is essential to the effec‐tive protection and management of these alpine ecosys‐tems(Wanget al.,2018).
Picea crassifolia(a species of cold-climate coni‐fer)is one of the most representative and dominant tree species in the forests of mountainous regions,es‐pecially in the Qilian Mountains(Changet al.,2014).Because the distribution of forests in the mountains appears to follow a pattern congruent with the topog‐raphy(Liuet al.,2013),it is commonly accepted that superior water conditions are created by the effects of the topography on solar radiation,leading to a more suitable habitat forPicea crassifoliagrowth on shady slopes(Kanget al.,2005;Nanet al.,2010).However,there is uncertainty about exactly how, and how much,various vegetation types affect soil-moisture content.For example,less radiation received in the forest on shady slopes may reduce the soil-water loss rate through evapotranspiration(ET);however,higher canopy interception can also reduce the soil-water in‐come from rainfall(Yanget al.,2017;Tianet al.,2018).In fact,one of our long-term field observations indicated that although the potential evapotranspira‐tion(PET)was significantly lower at the forest site we studied,the actual soil-moisture level under the forest cover was not as high as expected(Nanet al.,2010).Hence,these commonly held beliefs may not necessarily be true;and it is worthwhile to evaluate the soil-moisture and tree-water status for the forest under the ever-changing climatic conditions when ex‐ploring the ecohydrological mechanisms for vegeta‐tion patterns.
Indeed,soil moisture affects and links most of the fundamental processes acting in hydrological cycles(i.e.,evapotranspiration,deep percolation,and runoff(Laioet al.,2009))and ecosystem evolution(i.e.,veg‐etation patterning,adaptation,and distribution;(Ro‐driguez-Iturbe and Porporato, 2005)). Many tech‐niques,including time-domain reflectrometry(TDR),are available to automatically measure soil moisture in situ(Liuet al.,2015).Sap flow is a suitable indica‐tor of tree-water status under various weather condi‐tions(Gartneret al.,2009).It also can be measured automatically to track trees'physiological responses to environmental stresses(Buchmann,2002)and pro‐vide accurate estimates of tree-water supply and crown transpiration(Nunnet al.,2010).However,both soil-moisture dynamics and tree-water status are governed by nonlinear interactions among climate,soil,and vegetation(Manfreda and Fiorentino,2008);and many complex processes and uncertain variables are involved.Even long-term measurements are not likely to reveal a complete picture of soil-water dy‐namics or tree-water status;thus,a stochastic descrip‐tion is essential for capturing such nonlinearity,com‐plexity,and uncertainty(Laioet al.,2001;Porporatoet al.,2001).Soil moisture,as a stochastic variable,could lead to more probabilistic predictions of the car‐bon and water fluxes at a site(Milleret al.,2007).The stochastic method proposed by Rodriguez-Iturbeet al.(1999)and Laioet al.(2001)is one of the most widely cited frameworks.Starting from this probabi‐listic description of soil-moisture dynamics,Porpora‐toet al.(2001)further developed a set of analytical tools characterizing the plant-water status in waterlimited environments.
We hypothesized that both water and energy are limited for thePicea crassifoliaforest in the Qilian Mountains;further,a trade-off between reduced ET from limited solar energy and increased sustainabili‐ty of soil moisture may create a stressed but tolera‐ble environment and,in turn,produce an ecological niche relatively constant and favorable to the spe‐cies.As a first step toward examining the hypothe‐sis,the tree-water status of aPicea crassifoliaforest under its natural conditions was estimated by mea‐suring sap flow in a forest in the Qilian Mountains.We sought to identify the functional relationships be‐tween tree transpiration and environmental factors in the unique habitat by addressing two questions:(1)Is any water stress experienced during the growing season?and(2)What is the relationship between sap flow and soil moisture in the forest?Based on the re‐lationship established between sap flow and soil moisture,we further parameterized and tested the stochastic model of soil moisture,described earlier,with field measurements conducted in thePicea crassifoliaforest in the Qilian Mountains.The proba‐bility density function(PDF)of the long-term soilmoisture dynamics was then estimated with the vali‐dated model and parameters,and the analytical tools developed by Porporatoet al.(2001)were employed to test the proposed hypotheses fully by connecting the tree-water status with soil-moisture dynamics.This effort also gives a statistical and qualitative de‐scription of the tree-water status in a more general fashion,for further application in future modeling of vegetation patterns(Van Wijk and Rodriguez-Iturbe,2002;Cayloret al.,2005;Scanlonet al.,2005).
2 Material and methods
2.1 Site description
The study was carried out on the shady slopes in Dayekou(DYK,Qilian Mountains)(Figure 1),a rela‐tively small drainage system with an area of about 69 km2in the upper reaches of the Heihe River Basin(HRB,the second largest inland river basin in North‐west China(Gaoet al.,2008)).The climate in this re‐gion is temperate(Liuet al.,2015).According to the measurements during the period of 1980-2016 in this region,the long-term mean annual temperature was about 0.5°C,with the lowest temperature being-12.9°C in January and the highest 12.2°C in July.The mean annual precipitation was about 368 mm,occurring mainly during the growing season from May to Octo‐ber(determined by the air-temperature threshold of 5°C).The pan evaporation was 1,052 mm,and the mean annual relative humidity was 60%;the average annual sunshine duration was about 1,892 hours,and the average value of daily radiation is about 110 w/m2(Heet al.,2018).Forests are widely distributed on the shady and half-shady slopes at an altitude ranging be‐tween 2,400 and 3,300 m in this region and are domi‐nated byPicea crassifolia,a species of conifer in the Pinaceae family.The species can grow up to 25-m tall and 60 cm in diameter at chest height,with greenish or brownish yellow branchlets,and radially inserted leaves(Zhaoet al.,2007).Detailed forest-structure characteristics in the study area are given in Table 1.Dasiphora fruticosa,Caragana jubata,andSalix gilashanicaare some of the dominant shrubs in the study area.Polygonum viviparum,Carex atrata,andStipa capillata Linn.are some of the dominant species in the shady-slope grassland;andReaumuria soongorica,Sympegma regelu,Ceratoides lateens,andAchnatherum splendensare some of the dominant species in the sunny-slope grassland.
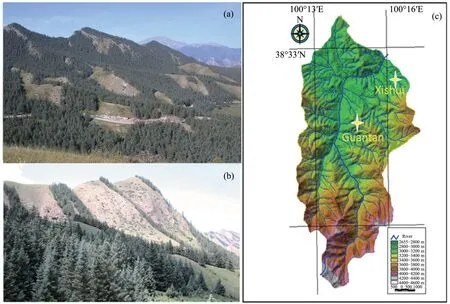
Figure 1 Self-organized alpine landscapes in arid Northwest China.The forests generally self-organize into ordered struc‐tures and coalesce into blocks on shady slopes((a),north-facing)or stripes along half-shady slopes((b),southeast-facing)in the altitude range of 2,600 to 3,200 m;and(c)location of the study(Dayekou catchment,modified from Yang et al.(2005))
The dominant soil textures are loam or light loam(gray-cinnamon soil)in the forests or grassland near the forest edges and loam or sandy loam(mountainchestnut soil)in the sunny-slope grassland.A typical profile of the shady forest soil consists of a graybrown A-horizon 7-to 8-cm thick and a light brown B-horizon 50-to 70-cm thick,underlain by a C-hori‐zon 30-to 50-cm thick.Soil profiles look quite differ‐ent in the sunny-slope grassland(light chestnut color),with a relatively thinner A-horizon(2-to 3-cm thick),a much thicker B-horizon(60-to 150-cm thick),and a variable C-horizon(30-to 190-cm thick).Root distri‐bution was plentiful in the A-horizon and upper B-ho‐rizon and sparse in the C-horizon(Wanget al.,2008).

Table 1 Structural characteristics of Picea crassifolia forest on south-facing shady slopes in the Dayekou watershed,Qilian Mountains
2.2 Soil moisture and meteorological measurements
Data used in this study were collected mainly from the Guantan Forest Station(100°15′0.8″E,38°32′1.3″N,2,835 m a.s.l.;Figure 1b.The observation site of Guan‐tan was equipped with an automatic system composed of sensors for measuring the microclimate of the for‐est.The measured factors and sensor types included net solar radiation(CM3 and CG3,Campbell Inc.,USA),air temperature and relative humidity(HMP45C,Vaisala,Finland),CO2(LI-7500,LI-COR Biosciences,USA),air pressure(CS105,Campbell Inc.,USA),soil temperature and moisture(CS107 and CS616,Camp‐bell Inc.,USA),geothermal flux(HFP01,Campbell Inc.,USA),wind velocity(014A,MetOne,USA),pre‐cipitation(52202,R.M.Young,USA),and eddy-covari‐ance ET(CSAT3 and LI-7500,Campbell Inc.,USA).Since 2007,measurements were recorded at 30-minute intervals by a datalogger(CR23XTD,Campbell Inc.,USA).More detailed information can be found in Tianet al.,2011.
The following measurements were included in the analysis:rainfall events,net solar radiation,eddy-co‐variance ET,and soil moisture,taken at depths of 5,10,20,40,80,and 120 cm by TDR sensors(the probes were installed sideways at the depths)and re‐ported at 30-minute intervals.To remove occasional gaps caused by failure of the power supply,weather data from the Guantan site were gap-filled with data from the nearby Xishui Forest Station(100°17′67″E,38°33′16″N,2,720 m a.s.l.,about 3.8 km from Guan‐tan).Volumetric soil moisture(θ)in the root zone(0 to 60 cm;Wanget al.,2008)was estimated with depth-av‐eraged soil moistures(10,20,and 40 cm)and then con‐verted to relative soil moistures usings=θ/n(nis the soil porosity).The following analysis uses daily aver‐ages of soil moisture and temperature,daily rainfall totals,and so on as shown in Figure 2.
2.3 Sap-flow measurements
Thermoelectric methods are the most widely used methods for determining sap flow of plants(Nadezh‐dina,1999;Roberts,2000).In this work,sap flow was measured in the trunks of three trees(Table 2,Figure 3)at the Guantan forest site during the grow‐ing season of 2008,May 1 to September 30,by using Granier's thermal dissipation systems.Generally,the system is based on the implantation of two needlelike probes(l=20 mm and φ=2 mm)and uses heat as a tracer of sap flow.The upper probe is continuously heated with a resistor,whereas the lower probe is un‐heated;the resulting temperature difference,ΔT,is measured with a thermocouple(Regalado and Ritter,2007).The probes were implanted into the xylem on the north side of each sample tree and connected to a datalogger to record ΔT(K).The ΔTcan be related to the rate of sap movement via the equation:

where ΔTmaxis the temperature difference between the heated needle and the sapwood ambient temperature when sap flow is 0,which is usually assumed to take place during the nighttime or predawn hours(Granier,1987).The SF-L sensor(UP GmbH,Germany)that we used in this study is an advanced version of the thermal dissipation system.It uses two more reference thermo‐couples to record background temperature gradients(ΔTr)of the sapwood,to correct ΔT,resulting in a much more accurate estimation of sap flow(Liu and Schweighoefer,2011).Because we did not use sap flow to estimate actual ET in this study,to simplify,ΔTmaxis assumed here to be equal to the maximum val‐ue of the calibrated ΔTfor every day(Granier,1987).An improved method of determining ΔTmaxcan be found in Oishiet al.(2008).For the same reason,we did not scale up the individual tree's sap-flow measure‐ments to plot or stand level(Mackayet al.,2010).

Figure 2 Time evolution of the sap-flow density and microclimate elements at the Guantan forest site during the growing season of 2008:(a)daily average sap-flow density and eddy-covariance evapotranspiration;(b)vapor-pressure deficit and net solar radiation;(c)relative soil moisture and rainfall;and(d)air and soil temperature

Table 2 Biometric and physiological parameters of sap-flow measurements
3 Data Treatment and Modeling
3.1 Statistical analysis
Evaluation of the potential relationships between the daily trunk transpiration(sap flow)and the various meteorological factors and soil moisture was per‐formed using linear correlation analysis on a daily time step for the entire growing season.Stepwise multiple linear regression analyses were performed to identify the key factors that affect daily transpiration for the se‐lected trees(the independent variables included net so‐lar radiation,vapor-pressure differentials,air tempera‐ture,relative humidity,and soil moisture in the root zone).All statistical analyses were performed with SAS version 9.3 software(SAS Institute),andp=0.05 was set as the threshold for statistical significance.
3.2 Soil-moisture model
The stochastic model of soil-moisture dynamics adopted here was originally developed by Rodriguez-Iturbeet al.(1999)and further modified by Laioet al.(2001)and includes both the probabilistic representa‐tion of the rainfall input and the state-dependent loss‐es for evapotranspiration and leakage.The model pro‐vides a realistic,quantitative description of the soilmoisture dynamics,while making the simplifications necessary to find an analytical solution.A detailed overview of the model can be found in Rodriguez-Iturbe and Porporato(2007).In the modeling frame‐work,lateral flow is not considered to affect soilmoisture dynamics;and the local recirculation of wa‐ter vapor is negligible.The foundation of the soilmoisture dynamics model is the water balance at a given point,using the equation:

where n is the soil porosity;s is the relative soil mois‐ture(the percent of the volumetric water content over the porosity or in the soil,over the sum of the volume of air and water);Zris the root-zone depth;Ris the rainfall depth;Iis the canopy interception;Eis the evapotranspiration(ET)rate;andLcombines deep in‐filtration and runoff losses(Laioet al.,2001;Rodri‐guez-Iturbe and Porporato,2007).The first two terms(R,I)represent the amount of infiltration into the root zone,and the last two terms(E,L)define the amount of water lost from the zone.
The occurrence of rainfall is idealized as a series of point events,arising according to a Poisson pro‐cess of rateλ,each carrying a random amount of rainfallh,extracted from an exponential distribution with meanα.Rainfall may be purged of canopy inter‐ception,which reduces both the frequency of effec‐tive rainfall and its depth;so the rainfall process is censored aswhere Δ is the rainfall depth threshold, below which no rainfall reaches the ground;and the mean rainfall depth(α)is reduced asα'=kα(kis the canopy throughfall coefficient)(Dalyet al.,2008;Vico and Porporato,2010).The sum of ET and leakage forms the soil-water-loss function(Milleret al.,2007).E[s,t]is modeled as a linear dependence of ET on soil moisture,from zero at the wilting point(sw)up to a maximum rateEmaxat the point of incipient stomatal closure(s*),and then con‐stant and equal toEmaxfor higher moisture levels.Deep infiltration and runoff losses,L[s,t],are as‐sumed to take place instantaneously whenever soil moisture reaches a thresholdsfc(soil capacity).When effective rainfall exceeds the available storage capac‐ity,nZr(sfc-s),any excess is immediately lost as runoff and deep infiltration(Rodriguez-Iturbe,Por‐porato,2007;Dalyet al.,2008).The analytical ex‐pressions for the steady-state PDF of soil moisture are given by Laioet al.,(2001).
3.3 Plant-water stress

where q is a measure of the nonlinearity of the effects of soil-moisture deficit on plant conditions.The mean value of water stress,˙(given that the plant is under stress),is calculated by considering only the part of the PDF corresponding toζvalues greater than 0(Por‐poratoet al.,2001;Porporatoet al.,2003).See Porpo‐ratoet al.(2001)for the detailed probabilistic struc‐tures and responsive performances ofζand
3.4 Parameter determinations
In this study,the soil-moisture dynamics model was validated in a deterministic mode for the forest site by using the real-time courses of rainfall as local forcing data.We used a reported relationship between the canopy-interception rate(I)and rainfall(R)to cal‐ibrate the rainfall input and estimate the rainfallthreshold Δ and the canopy-throughfall coefficient k(Tianet al.,2011):
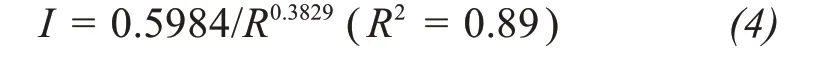
The value of the wilting point(sw)was calculated,from the linear relationship between soil moisture and sap flow,as the intercept of the regression lines with the horizontal axis.A double check was performed on the estimatedswby plotting the linear relationship be‐tween soil moisture and eddy-covariance ET.Field ca‐pacity(sfc)was determined by the difference between the dry weight of a soil sample and the weight after the sample had been moistened to excess and then allowed to drain for 24 hours;the point of incipient stomatal clo‐sure(s*)was assumed to be 80%of thesfc(Wanget al.,2008).Emaxwas assumed to be equal to the ET ats=s*,and was estimated from the linear relationship between actual ET and soil moisture(Figures 4a,4b).The pa‐rameters(λ,andα)characterizing the long-term precip‐itation were estimated directly from the rainfall records collected at nearby stations over the growing seasons of 1980 to 2010.Small calibration was applied to the mean depth of rainfall events(α)with a reported rainfallaltitude relationship for this region(Wanget al.,2008).The parameters and values are listed in Table 3.
4 Results
4.1 Measurement results
Climate.The year 2008 was relatively wet for the forest site of Guantan.Figures 2a-2d presents the sapflow dynamics,along with the key meteorological da‐ta,soil temperature,and soil moisture.In general,the growing season at the forest site was characterized by limited solar radiation,cool temperatures,frequent rain‐fall,and a relatively humid atmosphere even during the dry period.Mean solar radiation during the growing season was about 107 W/m2;and air temperature was on average 11.5°C,with a maximum daily average of 18°C.Wind speed was slow on a daily basis,peaking at 0.6 m/s.Sixty-one rainfall events with 357-mm cu‐mulative precipitation were recorded.Frequent rainfall(0.50/day),along with a weak wind(0.30 m/s),created relatively humid air masses(average 57.6%)and mod‐erate vapor-pressure deficit(VPD)(average 0.55 kPa).Dry days usually occurred after large rainfall events,allowing the VPD to increase and creating a relatively intense water demand in the trees,thereby creating strong evapotranspiration.

Table 3 Parameters of soil,plant,and climate characteristics at the forest site*
Soil temperature and moisture dynamics.Soil tem‐peratures of the root zone(60 cm)remained at a rela‐tively low level during the growing season(-0.1°C to 9°C for the surface layer and-1°C to 3°C for the lower layers),as did the soil-moisture storage(15%to 50%;Figures 2c,2d).At the beginning of the growing season,only 15%to 20%of liquid water remained in the root-zone soil;and this condition persisted until ear‐ly June.The frost in the soil was continually melting during the growing season and reached a maximum depth of about 110 cm in late September,as reflected in both the soil temperature and the moisture content of the profiles.Available liquid-water content in the rootzone soil was obviously controlled by the thermal bal‐ance before June 30;while after that,root-zone soilmoisture dynamics were more likely correlated with rainfall events of >15 mm and strong ET.The magni‐tude of soil-moisture pulses responding to rainfall was correlated with depth as expected(Figures 2c,2d).
Sap-flow dynamics.The first major increase of sap flow coincided well with the start of the growing season,May 1,and was then followed by a long peri‐od of a decreasing trend(until June 15).Soil moisture during this period was relatively low and stable and seemed not to exert a significant effect on daily sapflow changes.From early July until the end of the growing season,the major trend of sap flow was more likely to change with the moisture content of the rootzone soil,while solar radiation exerted a significant control over the day-to-day course(Figures 2a,2c).At the daily time scale,sap flow appeared to follow a diurnal trend with incident solar radiation.There was a pronounced diurnal hysteresis between the sap flow and solar radiation(-3 h between the peaks),which closely followed the course of VPD,especially on norain days.Typical diurnal courses of sap flow repre‐sentative of days with different weather conditions are shown in Figure 3.
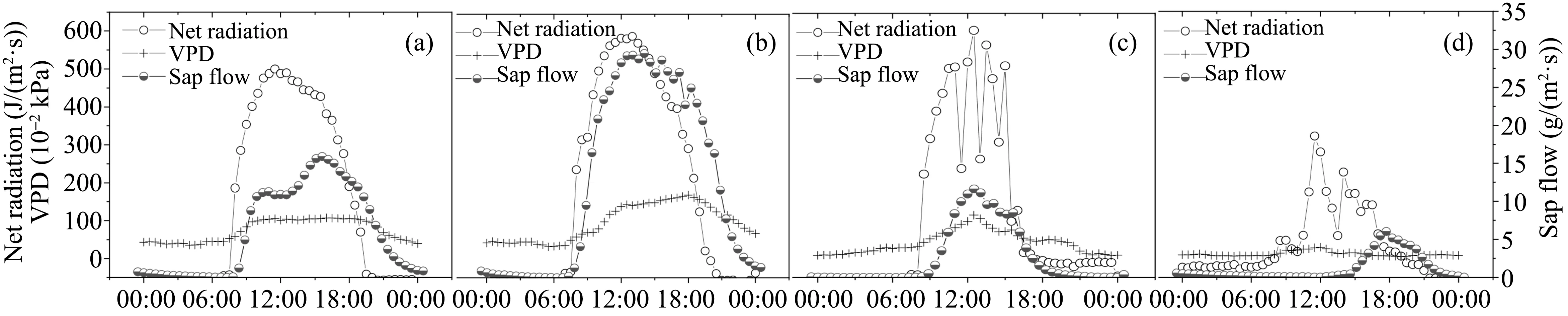
Figure 3 Hourly courses of sap flow,net radiation,and vapor-pressure deficit(VPD)on typical days of the growing season:(a,b)bright days(May 11 and July 5,2008,representing frost-affected and frost-unaffected days,respectively),(c)cloudy day(September 7,2008),and(d)rainy day(August 11,2008,8.6-mm precipitation)
4.2 Analysis and modeling results
Environmental control of sap flow.Changes in aver‐age daily sap flow in response to solar radiation over the growing season were quite apparent.Sap-flow rates were observed to increase with increasing net solar ra‐diation,without evidence of a saturation threshold dur‐ing the growing season of 2008(Figure 4a).A close correlation was also observed between sap flow and soil moisture,but it was greatly complicated by the frequent rainfall.After removing the rainfall days of the period from May 1 to July 30,2008,a relatively good,positive linear relationship(R2=0.74)between sap flux and measured soil moisture was obtained(Figure 4b).In addition,multiple linear regression analyses with stepwise backward procedures suggest that solar radiation and soil moisture were the most in‐fluential factors(explaining 43%and 26%of the vari‐ance in sap flow,respectively),followed by air tem‐perature,which could explain 4%of the variance in sap flow.The addition of other meteorological param‐eters(VPD,air humidity,and wind speed)enhanced the explainable variance by no more than 10%,attrib‐utable mainly to VPD.VPD appeared to be a less im‐portant driver of the seasonal course than it was of the diurnal course of tree transpiration(Figure 4c).

Figure 4 Relationship between daily sap-flow rate and(a)solar radiation,(b)soil moisture,and(c)vapor-pressure deficit(VPD)for the forest site during the entire growing season;piecewise fit curves for the open-circle data points are shown with the solid line
Modeling of soil moisture and water stress.We tested the stochastic soil-moisture model against data recorded during the period,starting from the begin‐ning of July and continuing until late September,for which soil moisture did not appear to have been af‐fected by either rainfall seasonality or the melting of frozen water in the root-zone soil.No-rainfall days(when soil moisture was drying out)during the period(2008)were selected to estimate parameters ofswandEmaxthrough the relationships between sap flow and soil moisture(Figure 5a)and between eddy-covari‐ance ET and soil moisture(Figure 5b).Almost the sameswvalues(0.20)were observed,and anEmaxof 4.4 mm/dwas determined from the relationships.Us‐ing the estimated parameters and time courses of rain‐fall as local forcing data,the stochastic soil-moisture model was validated in a deterministic mode for the forest site(e.g.,Kumagaiet al.,2009;Vico and Porpo‐rato,2010).Daily variations of calculated soil mois‐ture were compared against measurements for the pe‐riod of July 1 to September 30 for 2008 and 2010 in Figures 6a,6b),and the model-generated PDFs with actual daily rainfall were compared with the measured soil-moisture histogram in Figures 6c,6d).In general,the model reproduced measured soil-moisture content well,despite all the simplifying assumptions.The cal‐culated time series and obtained distributions provid‐ed a good fit of the observed soil-moisture time series and distribution in both the relatively wet(2008)and the dry(2010)growing seasons.Through the validat‐ed soil-moisture model and water-stress evaluation tools,PDFs of the long-term root-zone soil moisture and the static water stresses were numerically de‐rived,as shown in Figures 7a,7b.
5 Discussion
At the beginning phase of the growing season(May 1 to 30,2008),liquid soil moisture generally stayed below the estimated wilting point for the trees,although they continued to transpire,albeit at a mod‐erate rate.For example,the shape of the daily sapflow curve was typical of moderately water-stressed conditions on May 11(a bell-shaped curve,with the peak value of 11.4 g/(m2·s),Figure 3a),while the liq‐uid soil moisture was only 0.16,below even the wilt‐ing point of 0.20(Stöhr and Lösch,2004).A possible explanation may be the melting frost in the top soil layers(Mellanderet al.,2006).The moisture made available by the soil's thawing may become an impor‐tant source for tree-water uptake.As a result of the slow thawing,liquid water was gradually released,which probably prevented the development of an in‐tense water deficit and allowed the increase of transpi‐ration when evaporative demand was strong during a long rainless span(Repoet al.,2007).The soil-mois‐ture dynamics are obviously depth-dependent;sudden soil-moisture increases were observed at the rootzone depth in the soil profile during this period,asso‐ciated with the thawing of soil layers—a common characteristic in seasonal permafrost regions(Lopezet al.,2010).It is important to note that although TDR sensors have a valid range of-25°C to 50°C in tem‐perature,measurement errors are still possible and could mask the"real"soil-water content due to a drop in the dielectric constant when soil water was freezing(Paltineanu and Starr,1997).

Figure 5 Relationships(a)between the sap flow and soil moisture,and(b)between the eddy-covariance ET and soil mois‐ture over the root-zone depth of 0-60 cm during the period of July 1 to September 30,2008(when soil moisture was drying out and there was no rainfall);piecewise fit curves are shown with the solid line;sw,s*,and sfc refer to the wilting point,the point of incipient stomatal closure,and the filed capacity,respectively

Figure 6 Daily precipitation(bars,right scale);measured(line)and calculated(circle)daily average soil-moisture content(left scale)for the period of July 1 to September 30 in(a)2008 and(b)2010.Comparison between measured and modeled soil moisture shows the corresponding cumulative distribution functions(CDFs),probability density functions(PDFs),and histograms of the data for the period of July 1 to September 30 in(c)2008 and(d)2010.The parameters used for soil characteristics were sw=0.19,s*=0.57,and sfc=0.71
It appears that at the forest site and,generally,on the lower boundary of the shady or half-shady slopes of the Qilian Mountains,both water and energy are limited.We noticed that soil moisture in the root-zone depth responded substantially to rainfall events during this period,but the soil moisture was relatively low during the growing season overall(Figure 2c).The water status ofPicea crassifoliaas indicated by sap flow significantly responded to moisture variations over successive wet and dry spells,(Figures 2a,2c)and 4b).The estimated plant-water stress point,s*,was compared to the plots of soil moisture(Figure 5),indicating that the trees at the forest site spent almost all of the period from July 1 to September 30(during which root-zone soil moisture was assumed to be un‐affected by frost melting)under water stress(s*),nev‐er dropping below the wilting point(sw)except in the surface soil layers.A good linear relationship between sap flow and measured soil moisture in Figure 5 fur‐ther suggests strong evidence of soil-moisture depen‐dence(e.g.,Porporatoet al.,2001).This result is not surprising,especially when considering the limited rainfall and semiarid climate in the area(Porporatoet al.,2003).

Figure 7 (a)Estimated PDFs of the long-term soil moisture s and(b)static water stress ζ for the forest site(the atoms of probability at ζ=0 and ζ=1 are 1.5 and 0.6,respectively).Parameters of the graphs are α=6.7,λ=0.51,Emax=4.4,and q=3
Furthermore,changes in sap-flow rate were close‐ly related to solar energy input.The similar day-today variance and diurnal courses of sap flow and solar radiation suggest an intensive dependence of tree tran‐spiration on available solar energy.Multiple linear re‐gression analysis indicated that the energy factor ac‐counted for a considerable part of the variation within the data set,suggesting that high light intensity can in‐crease the sap flow inPicea crassifolia,given that wa‐ter is not limited.It is interesting that although photo‐synthetic and stomatal responses are generally consid‐ered to follow a hyperbolic function with light intensi‐ty(Gordonet al.,1999),sap-flow rates were observed to increase with increasing net solar radiation without evidence of an obvious saturation threshold.The ab‐sence of a saturation threshold may be attributed to in‐sufficient solar energy availability at the forest site,probably due to climate and topography(Lianget al.,2006).Limited solar irradiance could be the primary environmental constraint on vegetation activity;how‐ever,it could also lower the water on demand(Kum‐agaiet al.,2009).For example,the daily net solar ra‐diation and air temperature were only about 107 W/m2and 11.5°C,respectively,on average;in addition,a very low average ET rate of 1.32 mm/d(including soil evaporation,canopy transpiration,and canopy-in‐terception loss;Figure 2a)was measured at the site during the growing season of 2008(Tianet al.,2011).The reduced ET may contribute to lowering the water stress at the cost of lower growth rates;thus,an ET rate of 1.32 mm per day is not that low forPicea crassifolia,whose leaves have long life spans and thus generally low rates of gas exchange(Reichet al.,1999).We noted that the correlation of sap flow with VPD was unexpectedly weak,although this result was not surprising:Daily sap flux usually responds to wa‐ter and light first,and VPD explains variations after those variables are taken into consideration.Because solar radiation is highly correlated with temperature and VPD,an environment with high solar radiation contributes to warm temperatures and increased VPD;thus,multicollinearity of the variables could be one of the most likely reasons for this phenomenon(Gra‐ham,2003).Further,it could be partly related to the weak stomatal control of sap-flow rate likely caused by limited energy availability(Stöhr and Lösch,2004)or by canopy decoupling from low VPD and wind speeds(Figure 4)(Jarvis and McNaughton,1986;Jones,1992).For instance,low temperature and solar radiation may limit the growth rate and thus evaporative demand.Low VPD and wind speeds may not have triggered suf‐ficiently low leaf-water potential to cause stomatal clo‐sure in response to evaporative demand.Because the sap flow responded to VPD in a very nonlinear manner(Figure 4c),although we estimated the correlation be‐tween them through a linear regression analysis,this procedure also likely underestimated the VPD control of sap flow to some degree.
The stochastic soil-moisture dynamics model we tested captures the system's critical behaviors and cor‐rectly depicts the general shape of the histogram in both the relatively wet(2008)and dry(2010)growing seasons,in terms of the location(degree of saturation)and height (normalized frequency) of the peaks.Through the validated stochastic model of soil-mois‐ture dynamics,we numerically generated the long-term soil-moisture PDF(without considering the effects of seasonal frost),which is relatively open,with middle to low average values(Figure 7a)typical for such a cold region with a semiarid climate(Rodriguez-Iturbeet al.,1999).The calculated density of the static tree-water stresses(ζ)at the forest site is largely concentrated be‐tween 0 and 0.6(Figure 7b),suggesting a moderate wa‐ter-stress situation in most cases (Porporatoet al.,2001).This finding is sound evidence of a stressed but tolerable environment,probably created by a trade-off between reduced ET caused by limited solar energy and increased sustainability of soil moisture.The tradeoff results in a relatively constant ecological niche fa‐vorable toPicea crassifoliagrowth. The validated model can be used in further applications in studying and modeling vegetation-patterning processes,produc‐ing a more accurate explanation for the self-organized forest patterns in this mountainous region and thereby enabling the development of better management meth‐ods for their long-term sustainability.
6 Conclusions
This work investigated the soil-moisture dynamics and tree-water status in a forest site(Guantan)on the shady and half-shady slopes of the Qilian Mountains,where both water and energy are potentially limiting factors for vegetation.The functional relationships be‐tween tree transpiration and environmental factors were evaluated based on measurements of meteoro‐logical parameters and sap flow conducted in thePicea crassifoliaforest.Soil moisture and solar energy were identified as the most influential factors,explain‐ing more than 70%of the variance in sap flow.A sto‐chastic model of soil-moisture dynamics was parame‐terized and tested against the recorded field data.The steady-state probability density functions(PDFs)of the long-term soil-moisture dynamics and static treewater stress were estimated with the validated model and parameters.We found that the numerically gener‐ated PDF of the long-term soil moisture was relative‐ly open,with middle to low average values typical for such a cold region with a semiarid climate.The calcu‐lated density of the static tree-water stresses at the for‐est site was largely concentrated between 0 and 0.6,suggesting a moderate water-stress situation in most cases.Our results support the hypothesis that a tradeoff between reduced ET caused by limited solar ener‐gy and increased sustainability of soil moisture results in a relatively constant ecological niche favorable toPicea crassifoliagrowth in the alpine ecosystem.
Acknowledgments:
This research was jointly supported by the West Light Foundation of the Chinese Academy of Sciences(awarded to Dr.Hu Liu in 2018)and the Youth Inno‐vation Promotion Association of the Chinese Acade‐my of Sciences(awarded to Dr.Hu Liu in 2016).The data set for the Guantan Forest Station was obtained from the"Environmental and Ecological Science Data Center for West China"(http://westdc.westgis.ac.cn).Special acknowledgement is given to Professor Amil‐care Porporato(Duke University and Princeton Uni‐versity)for his generous help and useful discussion in preparing the manuscript.
杂志排行

Sciences in Cold and Arid Regions的其它文章
- Culturable bacterial diversity in hypolithic and peripheral soils in the west of the Hexi Corridor desert and its influencing factors
- A case study on Landscape Component Niche based on Landscape Pattern Indices:Yanchi,Ningxia Province,China
- Aeolian processes on sandy desertification of an alpine meadow:A wind tunnel experiment
- Characteristics of permafrost degradation in Northeast China and its ecological effects:A review