不同地理种源麻疯树表型变异研究
2020-05-30郭国业韩学琴
郭国业,徐 莺,唐 琳,陈 放,韩学琴
(1.南阳师范学院生命科学与技术学院,河南南阳 473061;2.四川大学生命科学学院,成都 610064;3.云南省农业科学院热区生态农业研究所,云南元谋 651300)
麻疯树(Jatropha curcas L.),又名小油桐,为大戟科(Euphorbiaceae)麻疯树属(Jatropha L.)多年生落叶灌木或小乔木[1]。麻疯树原产热带美洲,现广泛分布于美洲、非洲和亚洲的热带和亚热带地区[2]。麻疯树在中国多呈野生或半野生,广泛分布于云南、四川、贵州、广西和海南等地区。以云南分布面积最广,数量最多,集中分布于云贵高原南缘的亚热带干热河谷地带[3]。麻疯树耐干旱贫瘠,具有较好的抗逆性,是生态改良和荒坡地绿化的首选优良树种[4-5]。从麻疯树的叶、种子和根的汁液中分离提取的萜类、香豆素类和生物碱等物质具有抑菌、杀虫和抗氧化等多方面的药理活性[6-7]。麻疯树种油含量高,是当前重点研发的生物能源植物,也是世界公认的优良生物能源树种[8]。
基于分子标记的研究表明,非洲和亚洲地区分布的麻疯树种质资源的遗传基础较为狭窄,资源群体的遗传多样性普遍不高[9-11]。由于麻疯树雌雄花比例小和抗寒性较差等原因造成其产量不高,甚至在极端条件下不能结果[12]。通过对生物学表型性状的研究,发现不同地区分布的麻疯树在生物学特性、果实产量和种子质量特性方面也存在较明显的变异[13-15]。当前,我国大部分地区分布的麻疯树植物未被驯化而处于野生或半野生状态,现有品种良莠不齐,缺乏可大面积种植推广的优质高产的品种资源,限制了麻疯树作为能源植物的开发和利用。
加强对我国麻疯树种质资源的调查、搜集和生物学表型性状的研究对选育优质、高产的麻疯树品种资源和促进遗传改良具有重要意义。本研究通过对不同地理种源麻疯树的表型性状进行测定分析,以期揭示麻疯树的表型性状变异特征,探讨麻疯树不同种源间表型性状可能存在的遗传多样性,为麻疯树种质资源的保护和利用、遗传改良和品种选育提供参考依据。
1 材料和方法
1.1 试验材料
试验所用麻疯树种质材料采自云南省农业科学院热区生态农业研究所热带能源作物种质资源圃,该苗圃基地位于云南省元谋县的金沙江干热河谷生态气候区。其地理生境位于东经101°35′~102°06′,北纬 25°23′~26°06′之间的南亚热带干热季风气候区,海拔1 088~1 167 m。选取在地理分布和生境具有典型代表性的66份不同种源麻疯树,选取植株的树龄一致且均为5年,水肥管理情况基本一致,且生长状况良好。66份麻疯树种质资源的来源和生境信息见表1,依据种源地理分布将其划分为23个地理组群。
1.2 表型性状测定
每种源地随机选取10株进行表型性状的数据测定,每个性状指标重复测定3次,计算其平均值。测定表型参数指标:地径、株高、冠幅(东西和南北)、分支数(一级和二级)、结果量、果重、种形(种长和种宽)、百粒重和种子出仁率(种仁质量/种子质量×100%)。对植株的营养性状指标如地径、株高、冠幅和分支数的测定于当年6月,即麻疯树植株的营养生长趋于稳定时进行,测量工具为卷尺(0.10 cm)和数显游标卡尺(0.01 mm)。测定工作均在云南省元谋县云南省农业科学院热区生态农业研究所热带能源作物种质资源圃开展,选择晴朗无风的天气进行。对麻疯树的生殖性状如结果量、果实和种子的测定于10月初,即在植株的果实成熟期进行,果色为黄褐色时为宜。将采集到的果实带回实验室,置于60℃烘箱条件下烘干至果实重量恒定为止,测定果重、种形和百粒重等。
1.3 数据分析
利用Excel 2010对种质资源的表型数据进行均值、标准差和变异系数等分析。利用SPSS 12.0数据统计分析软件首先对表型数据进行标准化处理,在进行t检验和方差分析之前,首先对所有群组的性状数据进行正态检验以及方差齐性检验,检验结果发现所有数据均呈现显著性Significance(P值均大于0.001),服从正态分布假设,符合正态分布和方差齐性,可以进行方差分析(ANOVA)。然后对数据进行单因素方差分析、主成分分析和因子分析,基于欧氏距离(euclidean distance)和组间连接法(between-groups linkage)进行层次聚类分析(hierarchical cluster)。

表1 麻疯树种质资源和地理分布信息Table 1 Jatropha curcas germplasm resources and their geographical distribution information
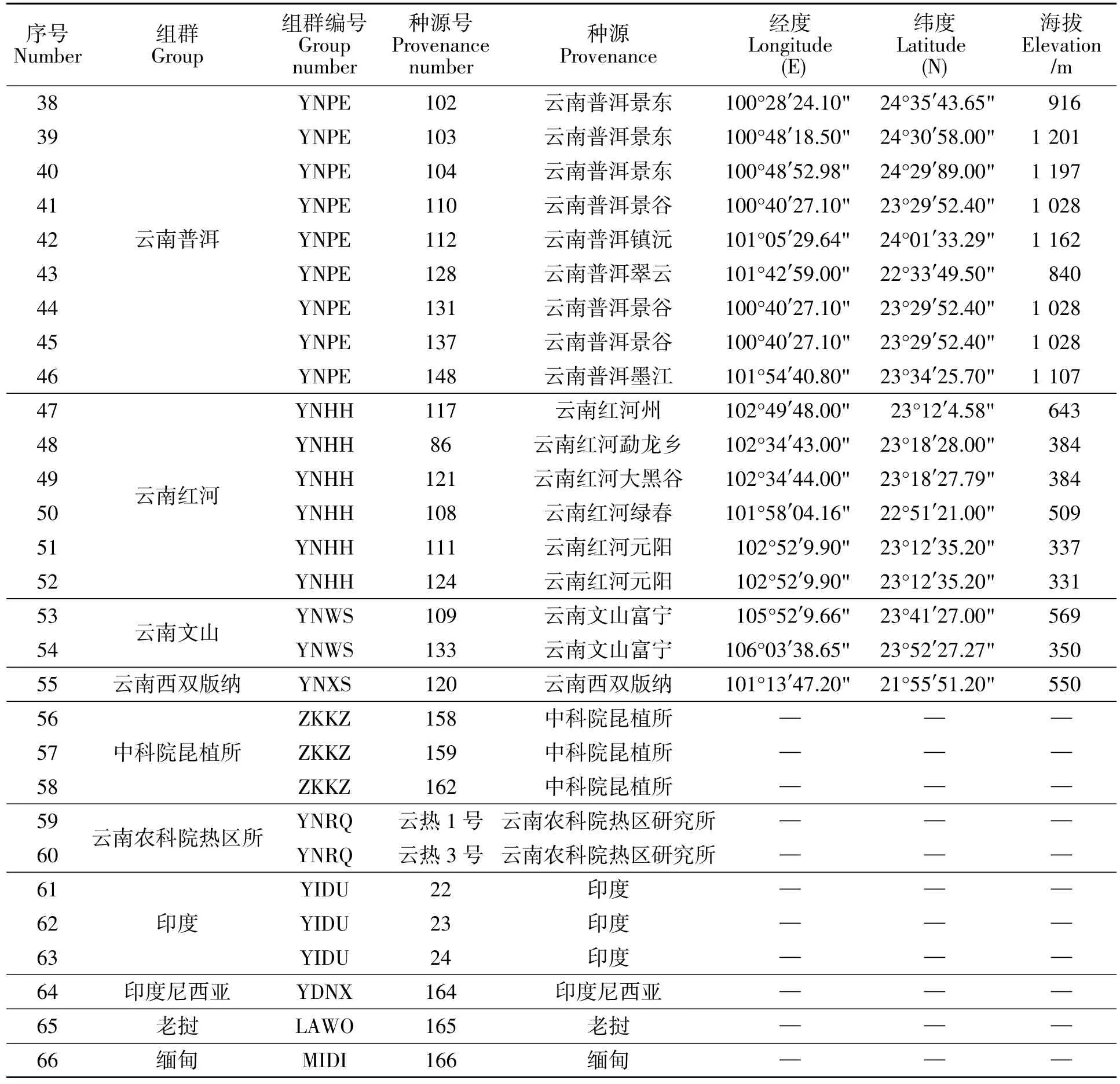
续表1
2 结果和分析
2.1 麻疯树种源组群间表型多样性
对23个麻疯树种源组群的地径、株高、冠幅、分支数、每株果簇数、每簇结果数、单果重、种长、种宽、百粒重和出仁率共11个植物学表型性状进行单因子方差分析(表2)。结果显示,不同地理种源组群在地径、株高、冠幅、分支数、每株果簇数、每簇结果数、单果重、种长、种宽、百粒重和出仁率方面在0.05的水平上均存在一定的差异性。其中,植株地径在各组群的性状间的差异呈显著水平(P<0.01),由中科院昆植所种源引种的植株地径最大(12.94±3.14),其次为云南楚雄(YNCD)种源(12.70±0.01),地径最小为云南昭通种源(5.75±1.43)。广西崇左种源的株高表现最高(3.53±0.25),其次为印度种源(3.36±0.28),云南玉溪种源的株高为最小(1.89±0.62)。综合冠幅(东西和南北)数据,四川攀枝花种源植株在冠幅方面较为突出(冠幅东西3.43±0.87,冠幅南北3.08±0.69),云南文山种源的冠幅为最低。二级分支数在各组群的性状间呈极显著差异(P<0.001),印度种源植株在分支数(一级分支数,二级分支数)上占有相对优势(5.00±1.00,13.33±1.15),其次为云南省农科院热区研究所选育品种(4.50±0.71,11±1.41),云南玉溪种源表现为最小(3.50±0.71)。云南文山种源的每株果簇数表现较高水平(141.00±5.66),其次为缅甸种源(121.50±7.78),云南玉溪种源的果簇数为最小(41.50±3.54);每簇结果数较高的为四川凉山(2.79±0.95),其次为由中科院昆明植物所引种品种(2.73±0.43),最低为云南曲靖(1.71±0.22)。印度种源在植株单果重方面具有较明显优势(2.66±0.19),其次为广西崇左种源(2.51±0.01),云南曲靖最低(1.45±0.06)。在麻疯树种实方面,种形(种长和种宽)、百粒重和出仁率在组群间的差异均不显著,种形较大的为云南玉溪(19.17±0.10,12.24±0.12),其次为云南楚雄双柏(18.45±0.58,11.16±0.16);百粒重较高的为印度种源(69.75±4.61),其次为四川凉山种源(66.49±1.54),最低为印度尼西亚种源(43.02±3.92);云南西双版纳、云南楚雄(YNCS)和四川攀枝花种源的麻疯树种质资源的出仁率具有相对明显的优势,印度尼西亚种源的出仁率表现最低。

表2 不同种源麻疯树组群的表型性状方差分析Table 2 Variance analysis of phenotypic traits between J.curcas groups among provenances
2.2 麻疯树组群表型变异分析
表型性状变异分析显示(表3),麻疯树表型性状间存在一定的差异。麻疯树营养生长的性状的平均变异系数较高,而生殖生长的性状的平均变异系数相对较小,不同麻疯树种源组群的表型性状变异依次为:二级分支数>一级分支数>每株果簇数>地径>每簇结果数>冠幅东西>单果重>百粒重>冠幅南北>株高>出仁率>种长>种宽。其中分支数、果簇数和地径的变异系数相对较高,表明这些表型性状存在遗传不稳定性;平均变异系数相对较小的为种形,即种宽(0.025)和种长(0.034),其次为出仁率(0.058),显示它们在麻疯树的表型性状中具有较稳定的遗传性。不同种源组群麻疯树的表型性状的平均变异系数也存在差异,变异系数较大的为云南普洱(YNPE)、云南红河(YNHH)、广西百色(GXBS)和云南楚雄(YNCY)种源的组群,其中,云南普洱(YNPE)组群的平均变异系数最大,为0.263,其次为云南红河(YNHH)为0.262,显示这两个地区麻疯树的表型多样性较高,遗传多样性较为丰富;平均变异系数相对较小的为云南曲靖(YNQJ)、老挝(LAWO)、缅甸(MIDI)和云南西双版纳(YNXS)种源组群,云南曲靖(YNQJ)组群的变异系数最小,为0.067,其次为老挝组群,为0.069,表明这两个群体的多样性相对较低。

续表2
2.3 麻疯树组群表型聚类分析
对23个麻疯树组群的11个表型性状进行主成分分析(表4)。结果显示,前4个主成分的累计贡献率达77.14%,可代表和保留原始因子中的绝大部分信息。第1主成分的方差贡献率为34.27%,起决定性作用的性状是百粒重(0.827),特征值为4.46。第2主成分对各变量的方差贡献率为18.93%,主要由种宽(0.703)决定,特征值为2.461,具有较高的正载荷。第3主成分对各变量的方差贡献率为13.14%,起决定性作用的性状是一级分支数(0.687)和二级分支数(0.681),特征值为1.708,具有较高的正载荷。第4主成分对各变量的方差贡献率为10.79%,起决定性作用的变量性状是一级分支数(0.536)。主成分分析表明麻疯树组群的表型多样性主要来源于百粒重、种宽、一级分支数和二级分支数4个性状。依据主成分分析确定的4个性状,采用欧氏距离法对23个麻疯树种源组群进行聚类分析(图1)。结果显示,在欧氏距离阈值M=10时,23个麻疯树组群被划分为5类群:第一类群是云南玉溪(YNYX)组群,主要特点表现为种长和种宽,即种形在组群间表现为最大;第二类群是印度(YIDU)组群,主要特点表现为二级分支数、单果重和百粒重在组群间最大;第三类群是云南昭通(YNZT)组群,主要表现为地径、种长和种宽在组群内为最小;第四类群包含印度尼西亚(YDNX)和云南农科院热区研究所(YNRQ)两个组群,印度尼西亚组群的冠幅(南北)、一级分支数为组群间最大,而百粒重和出仁率为最小,云南农科院热区研究所(YNRQ)种源性状表现为一级分支数和二级分支数较大,单果重为组群间最小;第五类群为多个不同地理种源组群交叉聚类,部分地理分布邻近的麻疯树组群聚类在一起。

表3 不同地区麻疯树组群的表型性状变异系数Table 3 Coefficient of variation of phenotypic traits in J.curcas groups from different distribution areas

表4 前4个主成分的负荷量、特征值、贡献率和累积贡献率Table 4 Factor loadings,eigen value,contribution rate and cumulative contribution rate of the first four principal components

续表4

图1 23个麻疯树组群表型性状聚类图Figure 1 Dendrogram constructed by Un-weight pair-group method using arithmetic averages(UPGMA)cluster based on phenotypic traits of 23 groups of J.curcas
3 讨论与结论
麻疯树在中国西南地区呈现野生和半野生分布,适生区为年均温20℃以上的南亚热带半湿润地区的金沙江、红河、澜沧江、南盘江和怒江流域,多生长于干热及湿热河谷地带[16]。不同种源来源以及西南地区复杂多变的气候和地理生态环境,使麻疯树的生物学性状呈现较明显的表型特征变异[17-18]。表型性状具有变异性和稳定性,受其本身的遗传组成和生态环境两方面的影响,是生物适应其生态环境的表现形式,表型变异越大,可能存在的遗传变异也越大[19]。本研究对66份不同种源的麻疯树种质资源的地径、株高、冠幅(东西和南北)、分支数(一级和二级)、每株果簇数、每簇结果数、单果重、种长、种宽、百粒重和出仁率共11个生物学性状的变幅和变异水平进行测定分析,显示各性状间均存在不同程度的变异。其中,分支数、地径和冠幅等以营养生长的植物学性状变异较大,而结果数、单果重、百粒重、出仁率和种形(种长和种宽)等以生殖生长的性状变异相对较低,显示麻疯树植物的营养器官对不同地理生境的反映更加敏感,更容易发生变异[19-20]。
本研究对植株地径、冠幅、每簇结果数、百粒重和出仁率等11个表型性状的方差分析显示,以云南楚雄、四川攀枝花和四川凉山等位于金沙江干热河谷分布区的麻疯树组群表现相对较高的变异水平。金沙江干热河谷相对高度差可达2 500 m以上,垂直分布差异显著,独特的地理气候可能是导致干热河谷的麻疯树分布自成体系而在表型性状呈现变异的重要原因[3,17]。不同种源组群麻疯树的表型性状的变异系数也存在差异,以云南普洱、云南红河、广西百色、云南元谋、云南丽江和四川攀枝花的麻疯树组群的表型变异相对较高,推测可能是由于地理隔离和有限的基因交流所造成。主成分分析表明,麻疯树组群的表型多样性主要来源于百粒重、种宽、一级分支数和二级分支数4个性状,依据4个性状参数对麻疯树组群进行形态聚类分析,成功将23个麻疯树种质组群划分为5大类群,各类群都能够依据形态特征的差异而分开,显示这4个性状是形成麻疯树表型变异的主要因子。沈俊岭等[21]对海南麻疯树试验基地2年生19个种源麻疯树的生长和结实性状研究,发现种群聚类未呈明显的地理相关性,认为中国境内麻疯树的变异为随机的地理变异模式。F.Maghuly等[22]综合利用AFLP、ISSR和SNP分子标记对收集于三个大洲15个国家共53个地区的麻疯树种质资源的群体遗传结构进行研究,发现亚洲和非洲麻疯树种质的遗传多样性相对狭窄,不支持其作为地理种源的真实起源,认为该地区麻疯树居群是由于人为强化选择导致的迅速扩张,而非自然野生分布。本研究综合聚类分析显示,部分同源地理组群能够聚类,但多数组群聚类呈现交叉,表明依据生长性状的组群聚类结果与种群的实际地理分布水平并不完全吻合,生长性状的组群划分与种源地理没有明显的相关性,显示麻疯树表型性状变异呈现不连续性,与前人研究结果基本一致。中美洲被认为是麻疯树的起源中心和多样性分布中心,推测不同的遗传背景、不同的迁徙扩散路径以及地理生境的差异可能是形成亚洲地区不同种源麻疯树组群表型性状多样性的主要原因[11]。
本研究通过对不同地理种源的麻疯树组群的形态学性状进行分析,发现不同种源间的地径、冠幅、分支数和每株果簇数的变异水平较大,暗示其容易受到环境压力的选择,而百粒重、出仁率、种实形状和种子出仁率的变异水平相对较低,在种质遗传进化中具有相对较高的稳定性。本研究分析并揭示了不同地理种源麻疯树群体的表型变异特征和变异来源,可为麻疯树的品种选育和种质创新等工作提供参考依据。在表型多样性分析的基础上结合分子标记技术进行遗传多样性评价将是今后研究的重要方向。
