X射线在棕榈藤纤维细胞壁结构研究上的应用
2020-05-29汪佑宏张菲菲季必超张利萍
汪佑宏,张菲菲,2,薛 夏,季必超,李 担,张利萍
1. 安徽农业大学林学与园林学院,安徽 合肥 230036 2. 黄山华塑新材料科技有限公司,安徽 黄山 245900
引 言
棕榈藤(rattan)属于棕榈科(Palmae)省藤亚科(Calamoideae),是具有重要经济价值和开发前景的热带和亚热带森林中的藤类植物资源。藤茎每年向上持续生长,具有多种用途,是仅次于木材和竹材的重要非木材林产品,为家具制作和工艺器具编制的优良材料[1]。目前,全球共有棕榈藤13个属664种,我国自然分布有4属37种,但经济价值较高的全球仅有20~30种[2]。
微纤丝角为细胞壁S2层微纤丝与细胞主轴之间的夹角。大量研究表明了利用X射线衍射法(XRD)能快速有效的测定式样的微纤丝角,具有重现性好、代表性强的特点,且测试速度快,在样本数量较大的变异研究中极其适用[3-4]。植物细胞壁主要是由纤维素、半纤维素和木质素组成,纤维素有结晶区和非结晶区(无定形区)交错联结而成; 纤维素的结晶度是指纤维素的结晶区质量(或体积)占纤维素整体质量(或体积)的百分数。利用X射线衍射法不仅能测定纤维素的结晶度,还有对其晶胞参数、晶粒尺寸等进行测试[5-7]。此外,X射线衍射法还可应用于微量物证鉴定[8]、表面残余应力[9]及木材含水率测试等。但微纤丝角和纤维素结晶度都能直接影响着材料的性能,是评价生物质材料的纤维品质和性质的重要依据,如微纤丝角小,其轴向干缩率小、顺纹强度大; 纤维结晶度高,则其吸湿性低、尺寸稳定性好等。本文通过对高地钩叶藤材微纤丝角、纤维素结晶度及微晶体尺寸及其变异进行研究,对于构建棕榈藤材细胞壁结构模型、探索棕榈藤强韧机理,为藤材高附加值加工利用提供理论依据。
1 实验部分
1.1 材料与设备
1.1.1 材料
高地钩叶藤(PlectocomiahimalayanaGriff)属钩叶藤属[1],丛生、攀援,采自云南南部的梁河县、海拔为1 480~1 500 m的山地常绿阔叶林中。藤径1.30~3.00 cm(2 m处),藤长16.0~20.0 m,节间长16.5~26.5 cm。分别在基部、2 m处、中部和梢部四个部位截取长约50 cm的节间、编号备用。
XD-3全自动多晶X射线衍射仪(北京普析仪器有限责任公司),Cu靶X光管,测角仪扫描半径为180 mm,扫描轴水平,测角仪角度重现性0.000 6°; X射线发生器kV、mA稳定度优于0.01%、额定功率3 kW。
1.2 方法
1.2.1 微纤丝角测试
分别在高地钩叶藤藤茎基部、2 m处、中部及梢部四个部位取样,再沿直径方向由一侧藤皮开始,依次、均匀选取尺寸为L(长)×T(厚)×W(宽)=25 mm×0.5 mm×W的薄片8片; 然后放置在温度为(20±2) ℃、相对湿度为65%±5%的调温调湿箱中平衡处理至少一周,使样品平衡含水率达到12%左右为止。
微纤丝角测定采用X射线衍射法,设定2θ=22.4°,测定范围70°~270°,测定速度为2°·min-1。
通常用(002)面的衍射强度曲线来求试样的平均微纤丝角,由于在测定范围内只有一个波峰,故选用0.4 T法计算该藤的微纤丝角,即40 %峰高处的宽度(M4)(见图1)。

图1 X射线衍射曲线(002)测微纤丝角Fig.1 Measuring MFA with X-ray diffraction curve(002)
1.2.2 结晶度及微晶体尺寸测试
在高地钩叶藤藤茎的基部、2 m处、中部和梢部分别选取藤皮、藤中和藤芯三个位置试样,使用球磨机磨成粉末后放入烘箱中在(103±2) ℃下烘至绝干。然后应用X射线衍射仪(Cu),在电压为36 kV,电流为20 mA,扫描范围为10°~45°,扫描速度为2°·min-1,采用逐步扫描法进行。
结晶度(CrI)按式(1)计算
(二)人民群众对音乐的欣赏需求差异性较大。我国地域广袤,人口众多,特别是在基层,人民群众在民族文化、传统风俗、地域特征、知识水平和艺术认知等各方面,存在着较大的差异性。而群文音乐面向的就是大众,需要具备普及性和易传播性,但在现实中,一个群文音乐作品在不同的群体中,受到的评价也是千差万别,甚至出现完全相反的评价,正所谓众口难调,这给群文音乐创作带来了极大的难度,也影响着创作者的信心和积极性。
(1)
式(1)中,CrI为结晶度(%);I002为002面晶格衍射角的极大强度;Iam为非晶格背景衍射的散射强度。
根据所得XRD谱图,还可以计算样品的微晶体尺寸等结晶参数(见图2)。其微晶体尺寸(L)可由Scherrer式(2)计算
(2)


图2 测定结晶度X射线曲线图Fig.2 Schematic diagram of measuringcrystallinity X-ray curve
2 结果与讨论
2.1 微纤丝角变异
2.1.1 微纤丝角径向变异
高地钩叶藤基部、2 m处、中部和梢部微纤丝角分别在22.53°~46.36°,30.06°~49.47°,29.14°~32.12°和23.47°~44.80°范围内变化,平均依次为34.94°,41.07°,35.02°和38.88°; 藤茎微纤丝角平均在29.93°~43.90°间变化,总平均值为36.50°,与黄藤材的36.1°相当、比钩叶藤材31.05°大17.55%。在藤茎上由藤皮-藤芯-藤皮方向,四个部位即不同高度上微纤丝角均呈抛物线的变异趋势,藤茎最外层的微纤丝角最小,而藤茎中间部位的微纤丝角最大[见图3(a)]。
由于微纤丝是组成细胞壁的基本单位,直接影响细胞壁的性质,从而影响材料的性质和用途,一般来讲微纤丝角越小,细胞的顺纹抗拉、抗压的强度越大,柔韧性越好,但伸缩性会变差,由此说明该藤藤茎最外层强度比藤芯好。
2.1.2 微纤丝角轴向变异
轴向上,对高地钩叶藤藤茎基部、2 m处、中部及梢部四个部位,分藤皮、藤中(位于藤皮与藤芯之间)和藤芯分别进行了微纤丝角的测试和分析。其中,藤皮处微纤丝角在26.52°~33.34°范围变化,梢部>2 m处>中部>基部; 藤中处微纤丝角在31.21°~36.17°范围变化,基部>2 m处>中部>梢部; 藤芯处微纤丝角在39.41°~48.97°范围变化,为2 m处>梢部>基部>中部。总体上藤茎微纤丝角在34.94°~41.07°范围变化,具体表现为2 m处>梢部>中部>基部,但规律性不强,可能原因是雨季和旱季形成的藤材(相当于木材的早、晚材)、藤龄等对微纤丝角有一定的影响,因取材时几乎无法保证全部取到早材或晚材,所以轴向变异规律性不强[见图3(b)]。

图3 微纤丝角径向(a)与轴向(b)变异Fig.3 The radial (a) and axial (b) variation of MFA
2.2 结晶度变异
高地钩叶藤藤茎基部、2 m处、中部和梢部纤维素结晶度分别在21.40%~34.01%,24.75%~36.45%,27.35%~35.58%和24.94%~34.60%范围内变化,平均依次为28.43%,31.18%,32.05%和29.98%; 藤茎纤维素结晶度平均在24.61%~35.16%间变异,总平均值为29.99%,分别比钩叶藤和黄藤高出29.57%和4.87%。
在轴向上,藤皮、藤中、藤芯及藤茎平均纤维素结晶度分别在34.01%~36.45%,25.37%~33.23%,21.40%~27.35%及28.43%~32.05%间变化,对应平均结晶度依次为35.16%,30.20%,24.61%和29.99%。藤茎平均纤维素结晶度从基部先逐渐上升,至中部达最大值后又下降,与藤中、藤芯处结晶度的轴向变化趋势完全一致; 而藤皮处结晶度也呈先上升后下降的变化趋势,但其最大值出现在2 m处。此外,除藤中处结晶度最小值出现在梢部外,藤茎平均、藤皮及藤芯处结晶度最小值均出现在基部(见图4)。
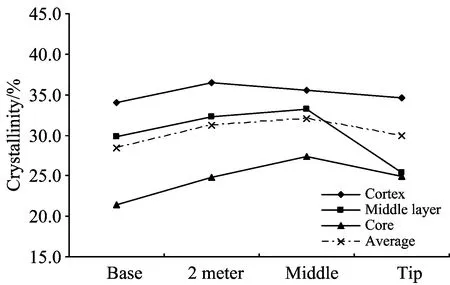
图4 结晶度的变异Fig.4 Variation of crystallinity
轴向上,藤茎纤维素结晶度为中部>2 m处>梢部>基部; 径向上,纤维素结晶度表现为藤皮>藤中>藤芯,且除梢部外,其余各处藤皮和藤中的结晶度明显要比藤芯高。因此,高地钩叶藤藤茎中段的大部分要比藤茎基部和梢部材质好、藤皮比藤芯质量高。
2.3 微晶体尺寸变异
2.3.1 微晶体宽度变异
高地钩叶藤材纤维素微晶体宽度在5.72~6.19 nm间变异,平均值为6.03 nm,是黄藤材微晶体宽度的2.51倍[10]; 其中藤皮、藤中、藤芯微晶体宽度分别在5.32~5.98,5.77~6.12,6.07~6.57 nm之间变化,平均微晶体宽度依次为5.74,6.02和6.33 nm。藤皮处微晶体宽度最小,藤芯最大。

图5 微晶体宽度(a)与微晶体长度(b)变异Fig.5 The variation of crystal width (a)and crystal length (b)
藤茎平均微晶体宽度与藤中、藤芯一样,随着藤茎高度的升高呈先下降后至中部达最小值后又上升的变化趋势,而藤皮处微晶体宽度呈“升-降-升”的趋势[见图5(a)]。
2.3.2 微晶体长度变异
高地钩叶藤纤维素微晶体长度在13.07~19.34 nm间变异,平均值为15.59 nm,是黄藤材微晶体长度的1.43倍[10]; 其中藤皮、藤中、藤芯微晶体长度分别在14.00~19.38m,13.02~18.31,10.61~20.32 nm之间变化,平均微晶体长度依次为16.17,15.28和15.31 nm。微晶体长度径向变化没有一定的规律; 而轴向随着藤茎高度的上升,均呈“降-升-降”的趋势,总体上微晶体的长度基部高于梢部,呈下降趋势,尤其是藤芯处,基部微晶体的长度比梢部高47.79%,这与黄藤的变化相反[10][见图5(b)]。
高地钩叶藤材抗弯强度(49.95 MPa)、基本密度(0.36 g·cm-3)等均小于黄藤材,高地钩叶藤材抗压强度(33.12 MPa)却大于黄藤材,而高地钩叶藤材纤维素微晶体宽度、长度均比黄藤材的大; 那么棕榈藤微晶体尺寸与其顺纹抗压强度、抗弯强度和基本密度等之间是否存在正、负相关关系,以及与其他物理、力学性质间存在一定的相关性,还有待进一步研究。
3 结 论
高地钩叶藤藤茎径向微纤丝角在29.93°~43.90°范围内变化,且藤皮处微纤丝角最小,藤芯处微纤丝角最大,说明藤皮强度比藤芯好; 轴向上藤茎微纤丝角在34.94°~41.07°范围变化,具体为2 m处>梢部>中部>基部,微纤丝角与藤龄间规律性不强。总体上藤茎微纤丝角在22.53°~49.47°间变异,均值为36.50°。
高地钩叶藤藤茎径向纤维素结晶度24.61%~35.16%间变化,径向上结晶度为藤皮>藤中>藤芯; 轴向上随藤龄的减小,结晶度呈先升后降变化趋势,且最大值在中部、最小值在基部。总体上藤茎结晶度在21.40%~36.45%间变异,平均值为29.99%。
高地钩叶藤材纤维素微晶体宽度在5.72~6.19 nm间变异,平均值为6.03 nm。藤皮处微晶体宽度最小,藤芯最大; 藤茎平均微晶体宽度与藤中、藤芯一样,随着藤茎高度的升高呈先下降后至中部达最小值后又上升的变化趋势。微晶体长度在13.07~19.34 nm间变异,平均值为15.59 nm。径向上微晶体长度为藤皮>藤芯>藤中; 微晶体长度轴向随着藤茎高度的上升,均呈“降~升—降”的趋势,总体上微晶体的长度基部高于梢部,呈下降趋势。
综合高地钩叶藤微纤丝角、结晶度分析,藤茎中段的大部分要比藤茎基部和梢部材质好、藤皮比藤芯质量高。
