红花瘤果茶种子贮藏特性及萌发过程中抗氧化酶活性的研究
2020-05-25郭米香1汪建文刘海燕3邹天才
郭米香1,洪 江,汪建文,刘海燕3,邹天才
(1.贵州大学林学院, 贵阳 550025; 2.贵州科学院,贵阳 550001; 3.贵州省植物园,贵阳 550004)
红花瘤果茶(CamelliarubituberculataChang ex Lin et Lu)为贵州特有珍稀濒危植物,散见于贵州省黔西南州喀斯特山区常绿落叶阔叶混交林亚乔木层和灌木层中,分布区狭窄、数量稀少[1-3],是山茶属植物进化发育保持了原始性状的类群,果实表面瘤状突起,木质坚硬,树形优美,颇具经济价值[4-5]。了解和掌握其种子的贮藏与萌发特性,保持种子活力,是开展好红花瘤果茶物种资源有效保护与合理利用的关键所在。影响种子贮藏寿命的因素包括内在因素和外在因素,其中内在因素包括种子生物学特性、含水量、成熟程度,外在因素主要为种子贮藏环境温度、湿度等[6]。籽粒饱满完整的种子萌发快,幼苗茁壮,贮藏寿命也较长。种子干瘪破损,其营养成分被破坏,不耐贮藏,容易腐烂。种子含水量及储藏的环境温度直接影响种子贮藏的活力质量,低温条件贮藏可能会延长或者缩短种子的寿命[6-8],同时,种子在萌发过程中的酶需要特定温度或光照的激活才能赋予其生理作用[9]。因此,研究种子贮藏的适宜条件及种子贮藏物质在贮藏过程中的生理特性或生化特征反应都可为种子的最佳贮藏方案提供科学参考,对珍稀濒危植物的繁育应用具有重要意义。
1 材料与方法
1.1 种子采集
2018年11月底,于贵州省晴隆县紫马乡龙头村上捧碧林区采集红花瘤果茶当年鲜种子,该林区为典型的高原亚热带湿润季风气候,岩石裸露60%~80%,海拔1 100~1 150 m,黑色石灰土,pH值7.9~8.2。种子千粒重1 466.60 g,分选除去瘪粒和虫害种子,置于通风干燥处备用。
1.2 种子贮藏
挑选出大小均匀的成熟种子,经1% KMnO4溶液消毒20 min并清洗,用布袋分装为5份,于2018年12月1日进行5种不同的贮藏处理。处理1:低温干燥处理,放于冰箱冷藏室(0~4 ℃);处理2:干燥冷冻贮藏(-24~-4 ℃);处理3:室内常温保存(15~25 ℃);处理4:拌沙冷藏,种子拌湿沙,放于冰箱冷藏室(0~4 ℃);处理5:拌沙冷冻,种子拌湿沙,放于冰箱冷冻室(-24~-4 ℃)。每30 d取样检测其种子脂肪、可溶性蛋白质、可溶性糖、含水量和生活力的变化,贮藏120 d后分别检测各组处理中的种子发芽率。未经贮藏的种子以25粒为1次重复,设置3个重复进行萌发试验[10],种子分别放置于15 ℃、25 ℃、35 ℃的温度条件下培养,保持每天10 h光照,并于25 ℃恒温下分别设置0 h、10 h、24 h等3个光照梯度,分别测定不同温度和光照处理下种子或幼苗的抗氧化酶活性。
1.3 生理指标检测
种子含水量采用加热烘干法测定,种子生活力采用TTC染色法测定[10];蛋白质含量采用考马斯亮蓝G-250染色法[9];可溶性糖含量采用蒽酮比色法,脂肪含量采用索氏提取法测量[10]。
1.4 种子发芽指标检测
种子萌发期间,统计种子发芽天数及其发芽数量,检测记录种子发芽动态,一直持续到种子萌发结束为止,3次重复后求平均值计算其发芽率和发芽势[10]。
1.5 种子萌发过程中抗氧化酶检测
未经贮藏的种子置于林木种子发芽盒中,在萌发过程中每间隔10 d取种子或幼苗,采用可见分光光度法检测过氧化物酶POD(试剂盒货号BC 0090;规格50 T/48 S)和超氧化物歧化酶SOD(试剂盒货号BC 0170;规格50 T/24 S),采用紫外分光光度法检测过氧化氢酶CAT(试剂盒货号BC 0200;规格50 T/48 S)。
1.6 实验数据
采用SPSS 21.0软件进行数据分析,采用Excel软件进行图表处理。
2 结果与分析
2.1 不同贮藏方法对红花瘤果茶种子含水量及生活力的影响
分别对5种贮藏方法的红花瘤果茶种子含水量、生活力进行检测,结果(图1)表明,在常温贮藏、干燥冷藏、干燥冷冻、拌沙冷冻4种储藏方式下,随着贮藏时间的增加,种子含水量逐渐降低,下降速率因不同贮藏方式而有差异。种子拌沙冷藏时,贮藏时间0~30 d,种子含水量开始升高,随后又逐渐下降,下降趋势不明显。红花瘤果茶种子在相同贮藏时间内,不同贮藏方式下的种子含水量存在差异。整体来看,拌沙冷藏的种子平均含水量均高于其他4种贮藏方式,常温贮藏条件下种子含水量最低。红花瘤果茶种子在贮藏过程中,种子生活力均呈下降的趋势,贮藏30 d、60 d、90 d时,生活力从高到低依次为拌沙冷藏、干燥冷藏、常温贮藏、干燥冷冻贮藏、拌沙冷冻贮藏。贮藏120 d后,种子生活力高低依次为拌沙冷藏、常温贮藏、干燥冷藏、干燥冷冻贮藏、拌沙冷冻贮藏。不同贮藏方式在相同时间节点下的种子生活力呈现出显著差异,其中拌沙冷藏处理的种子生活力在贮藏30 d、60 d、90 d、120 d均为最高。贮藏时间0~60 d时,种子生活力下降缓慢,贮藏60 d后,生活力开始急剧下降,甚至拌沙冷冻的红花瘤果茶种子在贮藏120 d后完全失去活力,种子生活力趋于0。拌沙冷藏条件下的种子生活力最高达30%,常温贮藏条件下种子生活力为20%,干燥冷藏及干燥冷冻种子生活力分别为18%、10%。
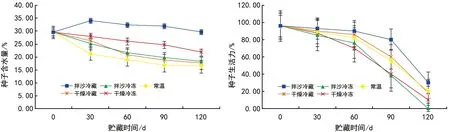
图1 不同贮藏方法对红花瘤果茶种子含水量及生活力的影响
2.2 不同贮藏方法对红花瘤果茶种子营养物质含量的影响
2.2.1不同贮藏方法对红花瘤果茶种子粗脂肪含量的影响
5种贮藏方式下的种子粗脂肪含量均呈下降的趋势(图2),可能是由于种子在贮藏过程中呼吸作用需要消耗脂肪来提供能量。不同贮藏方式下的种子粗脂肪含量由高到低依次为干燥冷藏(7.30%)、常温贮藏(6.56%)、干燥冷冻(6.34%)、拌沙冷藏(6.15%)、拌沙冷冻(6.01%),且在5种不同贮藏条件下,红花瘤果茶种子粗脂肪含量变化均存在差异。
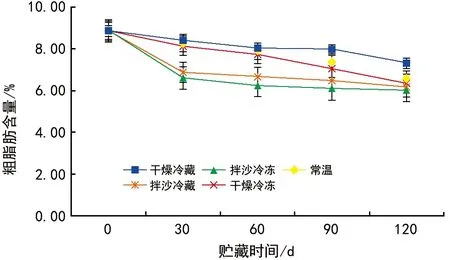
图2 不同贮藏方法对红花瘤果茶种子粗脂肪含量的影响
2.2.2不同贮藏方法对红花瘤果茶种子可溶性蛋白质含量的影响
不同储藏方法对红花瘤果茶种子可溶性蛋白质含量的影响如图3所示。整体上看,在不同贮藏过程中,可溶性蛋白质含量均呈现先升高后下降再升高的波浪式变化,其中可溶性蛋白质在0~30 d开始上升,随后30~90 d持续下降,在90~120 d又开始上升。其中,拌沙冷冻条件下贮藏0~30 d,可溶性蛋白质上升得最多,拌沙冷藏贮藏120 d后,可溶性蛋白质含量最高,拌沙冷冻的种子在贮藏120 d后可溶性蛋白质含量最低。红花瘤果茶种子贮藏30 d时,常温贮藏与拌沙冷藏条件下的种子可溶性蛋白质含量无显著差异,与干燥冷藏、干燥冷冻、拌沙冷冻贮藏条件下的种子可溶性蛋白质差异显著。贮藏60 d及120 d时,种子可溶性蛋白质均呈现差异,贮藏90 d时,干燥冷冻和拌沙冷冻贮藏条件下的种子可溶性蛋白质差异不显著,与常温贮藏、干燥冷藏及拌沙冷藏的差异显著。
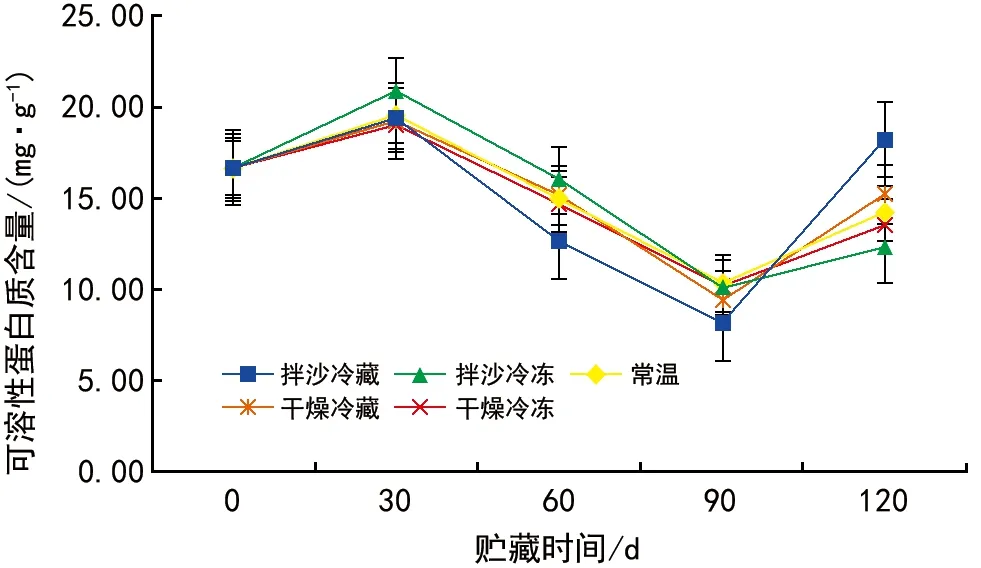
图3 不同贮藏方法对红花瘤果茶种子可溶性蛋白质含量的影响
2.2.3不同贮藏方法对红花瘤果茶种子可溶性糖含量的影响
不同贮藏方法对红花瘤果茶种子可溶性糖含量的影响如图4所示。随着贮藏时间的增加,常温贮藏、干燥冷藏、干燥冷冻、拌沙冷藏、拌沙冷冻条件下的红花瘤果茶种子可溶性糖均呈升高趋势。贮藏30 d时,常温贮藏与拌沙冷藏条件下种子可溶性糖含量差异不显著,干燥冷藏和拌沙冷冻的种子可溶性糖含量差异不显著。贮藏60 d,干燥冷藏和拌沙冷藏条件下的种子可溶性糖含量差异不显著,常温贮藏和拌沙冷冻贮藏可溶性糖含量差异不显著。贮藏90 d、120 d时,5种不同贮藏方式下的种子可溶性糖含量均呈现显著差异。
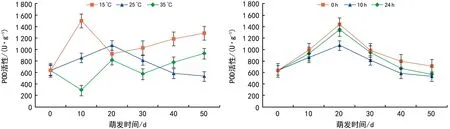
图5 温度和光照对红花瘤果茶种子萌发过程中POD活性的影响
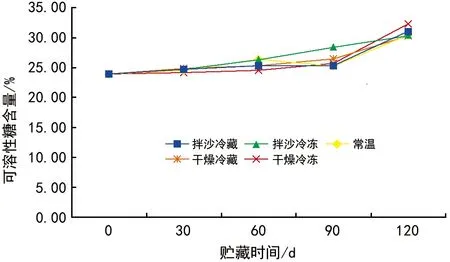
图4 不同贮藏方法对红花瘤果茶种子可溶性糖含量的影响
2.3 不同贮藏方法对红花瘤果茶种子发芽率和发芽势的影响
红花瘤果茶贮藏120 d后,于2019年4月5日进行萌发试验。结果(表1)表明,红花瘤果茶种子经过贮藏后的劣变程度较大,种子发芽率和发芽势降低,贮藏120 d后,种子生活力和含水量均下降,完好种子数量急剧减少。依次进行萌发试验,经拌沙冷藏120 d的种子含水量、生活力最高,初始发芽时间也较早,种子发芽率也最高,但远低于未经贮藏的种子发芽率。常温贮藏的红花瘤果茶种子发芽率、发芽势依次为8.00%、4.00%,干燥冷藏的种子发芽率和发芽势皆只为4.00%,主要原因是由于空气的干燥,种子呼吸作用促使种子营养物质消耗增加,剩余的营养物质不能支持种子的萌发。干燥冷冻的种子丧失了发芽能力,发芽率趋于0,拌沙冷冻贮藏的种子在贮藏结束后生活力也趋于0,其原因可能是冰冻导致细胞膜损伤,破坏了种子胚的活力而影响了种子的萌发。
表1 不同贮藏方法下的红花瘤果茶种子发芽基本情况
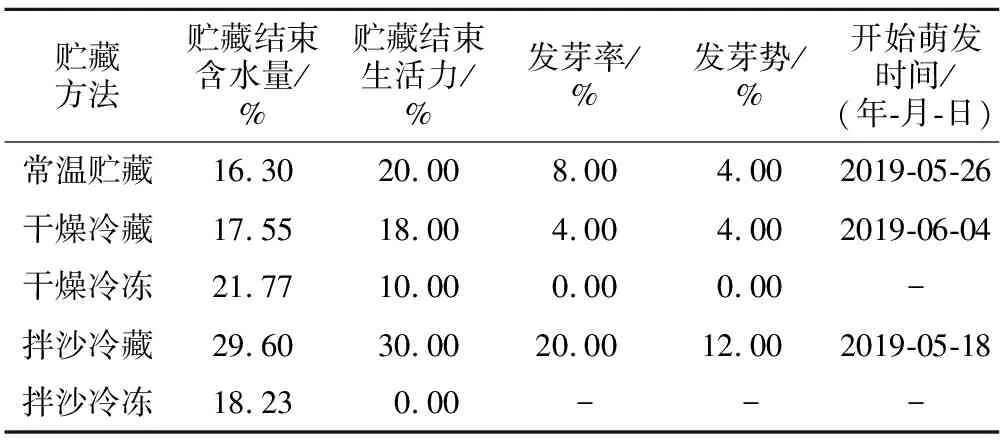
贮藏方法贮藏结束含水量/%贮藏结束生活力/%发芽率/%发芽势/%开始萌发时间/(年-月-日)常温贮藏16.3020.008.004.002019-05-26干燥冷藏17.5518.004.004.002019-06-04干燥冷冻21.7710.000.000.00-拌沙冷藏29.6030.0020.0012.002019-05-18拌沙冷冻18.230.00---
2.4 温度和光照对红花瘤果茶种子萌发过程中抗氧化酶活性的影响
2.4.1温度和光照对种子萌发过程中POD活性的影响
在不同的温度和光照条件下,测定种子萌发过程中POD活性的变化,结果如图5所示,25 ℃时,POD活性先升高后降低,在萌发0~10 d,POD活性上升,但上升速率远低于15 ℃条件下的,10~50 d,POD活性逐渐下降。在15 ℃条件下,POD活性在萌发0~10 d急剧上升,并达到一个高峰,随后在10~20 d急剧下降,在20~50 d又逐渐升高。35 ℃时,POD活性呈现先降低后升高再降低的波浪式变化,萌发初期0~10 d,POD活性下降到一个最低值,10~20 d又急剧上升,20~30 d开始下降,30~50 d又逐渐上升。总体来说,在15 ℃和35 ℃条件下,萌发后期的POD活性均高于萌发初期,高温和低温条件下,POD活性变化趋势较明显。当萌发温度一定时,不同的光照条件下,种子POD活性也呈现出相应的变化规律。其中0 h、10 h、24 h光照下的种子萌发过程中POD活性变化趋势一致,均为先升高,达到一个最高点后开始下降,POD活性的变化规律与光照长短的关系不明显。
2.4.2温度和光照对种子萌发过程中SOD活性的影响
SOD的活力与植物的抗逆性密切相关[20],在不同的温度和光照下对红花瘤果茶种子萌发过程中SOD活性进行了检测。结果(图6)表明,在15 ℃、25 ℃条件下,种子在萌发过程中的SOD活性呈现在0~20 d急剧下降,20~30 d上升,30~50 d又开始下降的波浪式变化。35 ℃下,SOD活性在0~10 d上升,10~50 d急剧下降。在15 ℃、25 ℃、35 ℃条件下均能发芽,说明SOD活性较强,种子内部生理活动旺盛,温度对种子SOD的活性存在影响。光照时间0 h、10 h、24 h条件下,SOD活性受到抑制,均在种子萌发0~20 d迅速下降,达到最低值,萌发20~30 d又开始上升,30~50 d再逐渐下降。不同光照条件下的种子SOD活性变化趋势一致,但在不同萌发阶段的SOD活性具有差异性,但该差异性远小于温度对SOD活性变化的影响。
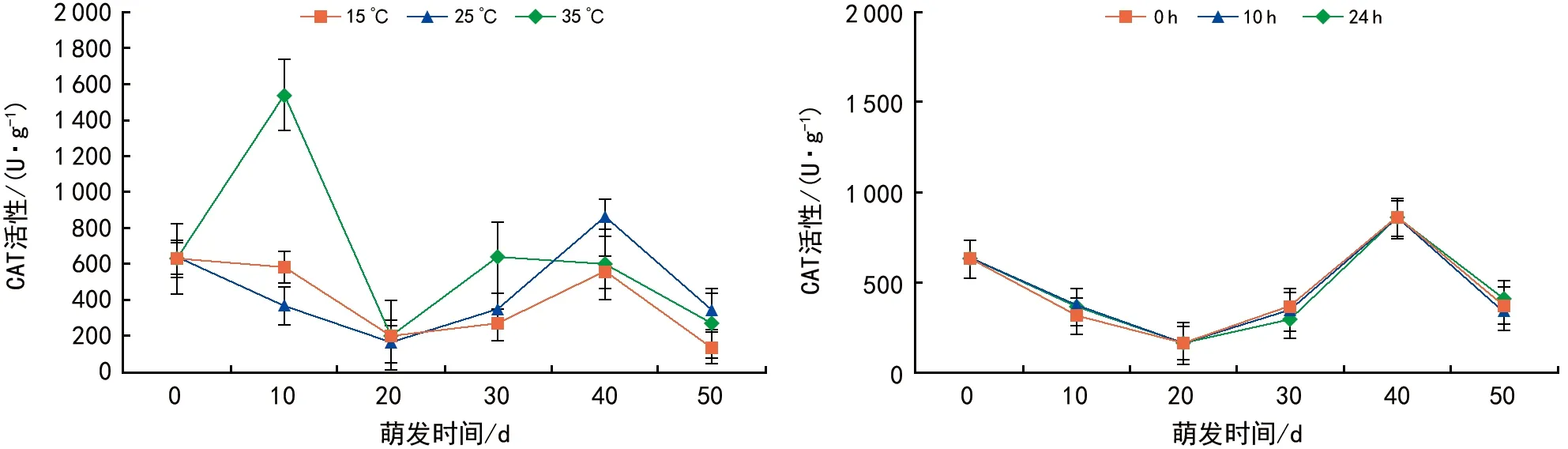
图7 温度和光照对红花瘤果茶种子萌发过程中CAT活性的影响
2.4.3温度和光照对种子萌发过程中CAT活性的影响
CAT能及时清除植物体内的有害H2O2,是植物体内主要酶促反应防御功能物质之一[21]。由图7可见,在3种不同的温度条件下,相同时间内种子CAT活性差异显著,种子CAT活性随着温度的不同,呈现出不同的变化规律。种子萌发0~10 d,35 ℃条件下的种子CAT活性最高,其次是15 ℃,25 ℃条件下CAT活性处于一个较低的水平,表明在种子萌发初期,过高或过低的温度都能使得CAT活性增强。种子萌发10~20 d,CAT活性下降,随后在20~40 d开始上升,最后40~50 d开始下降,萌发后期,种子发芽量增加,CAT活性下降。萌发40~50 d,25 ℃条件下的种子CAT活性高于15 ℃、35 ℃条件下的CAT活性,这也是红花瘤果茶种子在25 ℃条件下发芽率高于15 ℃和35 ℃的原因之一。在不同的光照时间下,红花瘤果茶种子CAT活性变化并未因光照时间不同而呈现明显差异,且在0 h、10 h、24 h光照下CAT活性变化趋势趋同,温度对红花瘤果茶种子萌发过程中的CAT活性的影响明显大于光照时间的影响。
3 结论与讨论
3.1红花瘤果茶种子适宜拌沙冷藏,种子活力有效贮藏时间小于120 d。拌沙冷藏的种子生活力最高,贮藏60 d时种子生活力达90.00%,贮藏60 d后种子生活力开始断崖式下降,在120 d时仅有30.00%,种子的含水量和生活力显著降低,当种子含水量下降到临界含水量以下时,会导致种子活力大幅度降低[11-12]。红花瘤果茶种子本身是高含水量种子,常温贮藏60 d后含水量开始下降,120 d后含水量降到最低(16.00%)。试验还表明,常温贮藏的红花瘤果茶种子发芽率为8.00%,干燥冷藏的种子发芽率仅有4.00%,干燥冷冻和拌沙冷冻贮藏的种子丧失了发芽能力,而拌沙冷藏120 d的红花瘤果茶种子含水量、生活力皆高于其他贮藏方式。因此,选择拌沙冷藏处理方式,合理调控种子贮藏时间等在红花瘤果茶种子育苗生产上具有推广应用的现实意义。
3.2红花瘤果茶种子内含的主要生化物质,在不同贮藏环境中随着贮藏时间的变化呈现出不同的规律性变化。研究种子贮藏过程中营养物质的变化有利于揭示种子在贮藏过程中的生理变化过程,可为探索种子贮藏方案提供科学依据[7],本试验5种贮藏方式条件下,红花瘤果茶种子粗脂肪含量均呈下降趋势,可溶性蛋白质含量均呈现先升高后下降再升高的波浪式反复规律,在贮藏时间为30~90 d时可溶性蛋白质含量急剧下降,贮藏90~120 d时可溶性蛋白质又开始升高,随着贮藏时间的增加,红花瘤果茶种子可溶性糖均呈现升高趋势。现代生物学研究表明,在种子贮藏过程中脂肪、可溶性蛋白质、可溶性糖之间可以相互转化,脂肪含量降低,相应的可溶性糖含量升高,糖是植物种子呼吸代谢的基质,可溶性糖含量升高[13-16];红花瘤果茶种子呼吸作用加强,种子加速其生理代谢,这将影响种子贮藏的质量和种子的生活力。
3.3不同温度和光照条件下,红花瘤果茶种子萌发过程中的氧化酶活性呈现出规律性变化。试验表明:
1) 较高或较低温度条件下,红花瘤果茶种子的POD活性变化趋势较明显。种子在较高或较低的温度条件下萌发,抗氧化酶促反应的活性降低,种子萌发到特定阶段,氧化酶活性又开始恢复并保持着生理平衡。
2) 红花瘤果茶种子在15、25、35 ℃条件下均能发芽,3个温度梯度条件下的SOD均有活性,在萌发后期,35 ℃条件下的SOD活性低于15 ℃、25 ℃,表明温度偏高将会导致SOD活性降低,从而影响种子的萌发出苗。
3) 红花瘤果茶种子CAT活性随着环境温度的不同,呈现出不同的变化规律。种子萌发0~10 d,35 ℃条件下的种子CAT活性最高,15 ℃次之、25 ℃再次之,说明在种子萌发初期,较高或者较低的温度都能使得CAT活性增强。但在萌发后期(40~50 d),25 ℃条件下的种子CAT活性明显高于15 ℃和35 ℃,也是红花瘤果茶种子在25 ℃条件下发芽率高于15 ℃和35 ℃的原因之一。
4) 在不同光照时间下,红花瘤果茶种子萌发过程中POD、SOD、CAT的变化趋势并未随着光照时间的变化而发生变化,光照时间的长短对红花瘤果茶种子萌发过程中的抗氧化酶变化的影响不明显。已有研究证实,当植物种子在萌发过程中受到环境胁迫时,产生的活性氧需要抗氧化系统来免疫清除,POD、SOD、CAT等发生生理功能作用,维持细胞自由基的低水平状态和保护细胞膜不受损伤,以维持细胞内活性氧处于一个较安全的水平[18-22]。红花瘤果茶的种子也不例外,研究掌握种子贮藏过程中的生理生化特性,探索其种子萌发过程中抗氧化酶变化规律等具有重要科学意义和生产应用价值。
