日本无刺花椒抗寒性的初步研究
2020-05-24李佩洪吴银明
陈 政,李佩洪,唐 伟,龚 霞,吴银明,吴 军
(四川省植物工程研究院,四川资中 641200)
花椒(Zanthoxylum bungeanum Maxim)原产于我国中西部,为芸香科花椒属落叶灌木或小乔木。全世界约有花椒属植物250 种,分布于亚洲、非洲、大洋洲、北美洲的热带和亚热带地区[1]。中国是世界花椒第一生产大国,有39 种14 变种[2]。四川是我国花椒主产区之一,汉源花椒、茂县大红袍花椒、金阳青花椒、洪雅藤椒、蓬溪青花椒是四川花椒中的五朵金花,是全国优质花椒之一。由于花椒树干、枝、叶具刺,在育苗、栽植、管理和采收等方面有诸多不便,尤其采收难度大、成本高,花椒采收难是花椒生产中亟待解决的关键问题,若能以无刺或少刺花椒品种替代有刺花椒,将会大幅度提高采收效率、节约采收成本。为解决生产中花椒刺多的问题,我院于2009 年从河北林业科学研究院引进花山椒、朝仓山椒、葡萄山椒和琉锦山椒等无刺花椒品种进行引种栽培试验。日本无刺花椒(Zanthoxylum Japonica),为落叶小灌木,雌雄异株,无刺或少刺。树势直立,萌芽力和成枝力均强。雌花具有雌蕊1~3 枚;雄花具有雄蕊5~6 枚,花粉量大。果实圆形或椭圆形,油腺点小且密,果皮鲜红色[3]。果皮、青果、嫩芽、种子等均有重要的应用价值。日本无刺花椒由于无刺或少刺,采摘和管理方便,较“茂汶花椒”“汉源花椒”“九叶青花椒”“藤椒”等花椒品种节约大量采摘用工和管理用工。日本无刺花椒在引种、育苗、栽培机理、修剪技术、施肥技术、风味物质及离体愈伤组织等方面做了大量工作[4-12],但对无刺花椒抗寒性研究报道较少,毕君[8]对1a 生枝条的细胞膜透性和SOD 活性变化、POD 活性变化、MDA含量变化、自然越冬的枝条电导率与冻害3 种途径进行抗寒评价,而红花椒抗寒性研究报道较多[13-17]。为此,笔者对无刺花椒材料的抗寒性进行初步研究。花椒抗寒性研究与农业生产密切相关。解决花椒寒害的途径很多,其中选用耐寒的优良花椒品种是最经济、也是最有效的途径之一。因此,探索花椒抗寒性生理机制,不仅在理论上有重要意义,而且在生产中也具有广泛的实用价值。
1 试验材料与方法
1.1 试验地概况
试验地设在四川省资中县明心寺镇,位于29°44.29′N,104°56.38′E,年均气温17.4℃,极端最低气温-3.2℃,极端最高气温41.9℃,年均日照时数1246.5h,年均降雨量1007.7mm,降雨量主要分布在5~9 月。土壤为砂壤土,pH 值7.0,海拔310m。
1.2 试验材料
测定的材料有朝仓山椒、葡萄山椒、琉锦山椒、花山椒,共4 个品种,树龄为2a。
1.2.1 朝仓山椒。枝条密集,成抱头状生长,新梢上部绿色下部为棕色;树皮光滑,基本无刺;叶片长宽均值为20.66mm/9.62mm,叶缘为锯齿状,向内卷曲,小叶数多为11~15 枚;果实圆形,果实纵横径均值为4.85mm/4.46mm,脐部有一小突起,有残留花柱,果皮鲜红色[18]。油腺点小、密而深陷。花期为3 月中下旬,果期为8 月中下旬。
1.2.2 葡萄山椒。枝条较疏张,生长势较弱;有皮刺;新梢上部绿色,下部为棕色;叶片长宽均值为24.66mm/12.28mm,叶缘为锯齿状,向内卷曲,小叶数多为9~17 枚;果实较大,果实纵横径均值为6.45mm/5.07mm,脐部有一小突起,有残留花柱,果皮鲜红色[18]。油腺点小且密而深陷,花期为3 月中下旬,果期为8 月中下旬。
1.2.3 琉璟山椒。树姿较舒张,生长势较强;无刺;新梢上部绿色,下部为棕色;叶片长宽均值为21.27mm/10.17mm,叶缘为锯齿状,叶片平整,小叶数多9~15 枚;果实较大,圆形,纵横径均值为5.94mm/5.01mm,脐部有一小突起,有残留花柱,果皮鲜红色[18]。油腺点小、密而深陷。花期3 月中下旬,果期为8 月下旬~9 月上旬。
1.2.4 花山椒。树势强,枝条粗壮;无刺;叶片长宽均值为25.67mm/12.26mm,叶缘为锯齿状,向内卷曲,小叶数多为13~15 枚;成花容易且花粉量极大,是比较理想的授粉品种[18]。花期为3 月上中旬。
1.3 试验方法
1.3.1 植物材料处理。2019 年4 月,取样时间为上午7:00~9:00,剪取健康、无病虫害的朝仓山椒、葡萄山椒、琉锦山椒、花山椒花椒幼树树冠中部外侧的4 个方位的夏梢顶部向下的第4 叶、第5 叶(完整叶),将叶片用洁净的湿纱布擦拭并晾干,按品种、处理分装于塑料袋中,放入冰箱内进行人工冷冻,处理温度分别为-6、-4、-2、0、2 和4℃并保持恒温5h。随后,将各处理叶片放入-20℃超低温冰箱中保存,用于测定SOD 活性、POD 活性、MDA 含量生理指标,设3 个重复。
2019 年1 月上旬在资中试验地内,选取朝仓山椒、葡萄山椒、琉锦山椒、花山椒4 种花椒的健康、无病虫害的1a 生枝条作为供试材料。将各花椒的枝条用洁净的湿纱布擦拭并晾干,放入冰箱内从0℃开始降温,降温速率为4℃/h,处理温度分别为:-4、-8、-12、-16、-20、-24、-28℃,并保持恒温12h。再以同样的速率升温至0℃,取出枝条用于REC 的测定,以放置室温下(约为0℃)的枝条为对照(CK),设3 个重复。
1.3.2 指标测定方法。过氧化物酶(POD)活性测定采用愈创木酚法,超氧化物歧化酶(SOD)活性测定采用NBT(氮蓝四唑)光化还原法,丙二醛(MDA)含量测定采用硫代巴比妥酸法,电解质外渗率采用电导率法。数据取各生理指标3 次测定结果的平均值。以上测定方法均参考文献[19]。
1.3.3 数据处理。数据处理用EXCEL 和DPS(v6.85 版)软件完成。
2 结果与分析
2.1 SOD 酶活性的变化
在正常条件下,植物细胞内自由基的产生与消除处于平衡状态,这样可以保持细胞膜系统的稳定性,维持正常的生理代谢活动。当植物处于各种逆境胁迫时,这种平衡会受到破坏而出现自由基的累积,从而引发或加剧细胞膜脂过氧化作用,造成膜系统受损,严重时导致细胞死亡。在低温逆境条件下,SOD 能被活性氧诱导产生,从而减轻对细胞膜的伤害[20]。SOD、POD 是植物体内清除自由基的主要酶类,其活性大小反映细胞对逆境的适应能力,也间接表明植物耐寒性的强弱。
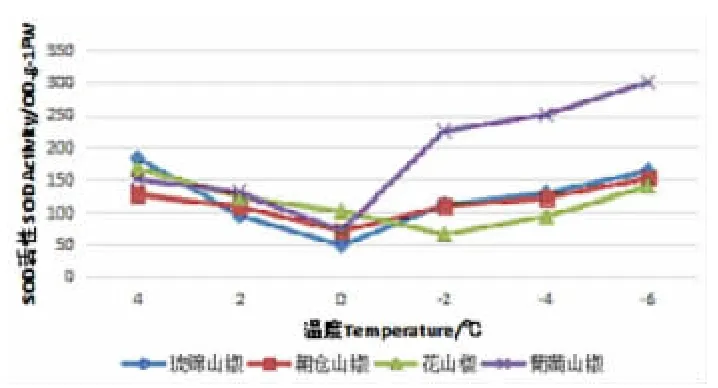
图1 降温处理过程中的SOD 活性变化(单位:u·gFW-1·h-1)
植物在低温胁迫初期,在植物细胞内,SOD 主要起清除超氧自由基的作用。图1 所示,各花椒SOD 活性随着低温胁迫的加强显示出了先下降后增加的变化趋势。随着低温的加深,4 个山椒品系都是从4℃降到0℃时SOD 活性下降,从0℃降到-6℃时上升,SOD 活性变化呈“V”趋势。SOD 活性上升速率并不均匀,在0℃降到-2℃时SOD 活性上升较快,其中葡萄山椒SOD 活性上升最快,琉锦山椒次之,朝仓山椒SOD 活性上升最慢。花山椒SOD 活性的变化趋势不同,花山椒4℃降到-2℃时SOD 活性下降,从-2℃降到-6℃时呈现上升趋势,活性变化呈“V”趋势。可见,从SOD 活性上升速率来看,葡萄山椒的上升速率最快,琉锦山椒的较快,朝仓山椒的次之,花山椒的最慢。
2.2 POD 酶活性的变化
过氧化物酶(POD)是植物体内普遍存在的活性较高的一种酶,它能够清除超氧自由基形成H2O2[20],从而保护植物免受活性氧伤害。图2 所示,4 个山椒品系POD 活性随着低温胁迫的加强显示出了花山椒先下降后增加后又下降再增加的变化趋势。随着低温的加深,都是从4℃降到2℃时POD 活性下降,从2℃降到0℃时上升,0℃降到-2℃时呈现下降趋势,从-2℃降到-6℃时又呈现上升趋势,活性变化呈“W”趋势。朝仓山椒、琉锦山椒和葡萄山椒从4℃降到-2℃时下降,从-2℃降到-6℃时上升,活性变化呈“V”趋势,POD活性上升速率不均匀,4 个山椒品系在-2℃降到-4℃时上升较慢,在-4℃降到-6℃时上升较快,葡萄山椒POD 活性上升速率最快。可见,从POD活性上升速率来看,葡萄山椒的最快,花山椒的较快,朝仓山椒的次之,琉锦山椒的最慢。
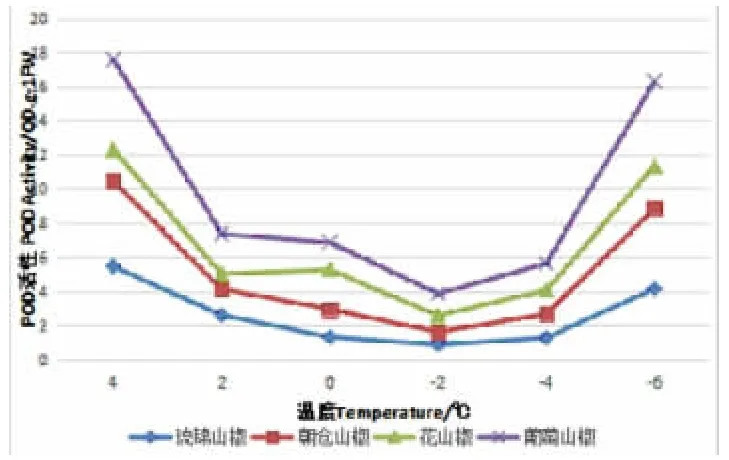
图2 降温处理过程中的POD 活性变化(单位:△OD470·gFW-1·min-1)
2.3 MDA 酶含量的变化
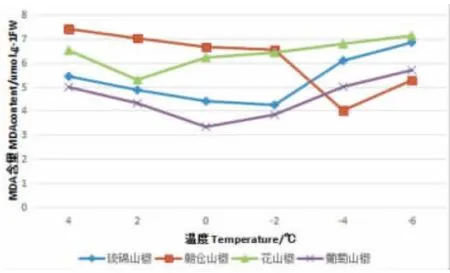
图3 降温处理过程中的MDA 含量变化(单位:μmol/g.FW)
植物在逆境条件下往往发生膜脂过氧化作用,其产物MDA 会严重损伤生物膜。通常利用它作为膜脂过氧化指标,表示细胞膜脂过氧化程度及对逆境反映的强弱,因此在研究植物寒害及抗寒性时,测定MDA含量的变化可作为一项生理鉴定指标[21]。从图3可知,4 个山椒品系MDA含量随着低温胁迫的加强大多显示出先下降后上升的趋势。各山椒品系MDA含量上升温度不同,增幅也不同。花山椒MDA 含量从2℃开始上升,2℃降到0℃时其MDA 含量上升了0.926μmol/g.FW,随后增幅变缓;葡萄山椒MDA 含量从0℃开始上升,0℃降到-2℃时其MDA 含量上升了0.504umol/g.FW,-2℃到-4℃时MDA 含量上升了1.162umol/g.FW,随后增幅变缓;琉锦山椒MDA 含量从-2℃开始上升,-2℃降到-4℃时其MDA 含量上升了1.843umol/g.FW,-4℃降到-6℃时其MDA 含量上升了0.754umol/g.FW;朝仓山椒MDA 含量从-4℃开始上升,-4℃降到-6℃时其MDA 含量上升了1.090umol/g.FW。可见,琉锦山椒MDA 含量增幅最大,葡萄山椒的次之,朝仓山椒的再次之,花山椒的最小。
2.4 不同低温处理对花椒枝条质膜透性的影响
朝仓山椒、葡萄山椒、琉锦山椒、花山椒4 个山椒品系1a 生枝条经低温处理后,其REC 如图4所 示。从图4 可以看出,在一系列冷冻低温处理下,4种花椒的REC 与处理温度之间关系呈上升的“S”形曲线。但其REC 增加的速率并不均匀,处理温度越低REC 变化相对越快,且因种类而异。当温度高于-16℃时,随温度降低,枝条的相对电导率缓慢上升,-16~-24℃期间,相对电导率急剧增加,之后趋向平缓。-16℃下降至-20℃时,4 个山椒品系相对电导率变化为:琉锦山椒的相对电导率增幅最大,为19.2%;花山椒的相对电导率增幅次之,为15.5%;葡萄山椒的为14.3%;朝仓山椒的相对电导率增幅最小,仅为7.9%。相对电导率急剧增大,说明枝条组织已经受到严重伤害,膜透性增加,细胞内电解质大量外渗。由此可知,在相同低温下,琉锦山椒电解质外渗最多,脂膜损伤程度最重;葡萄山椒与花山椒电解质外渗较少,朝仓山椒电解质外渗最少,脂膜损伤程度较轻,这表明不同花椒种类的抗寒性存在差别。
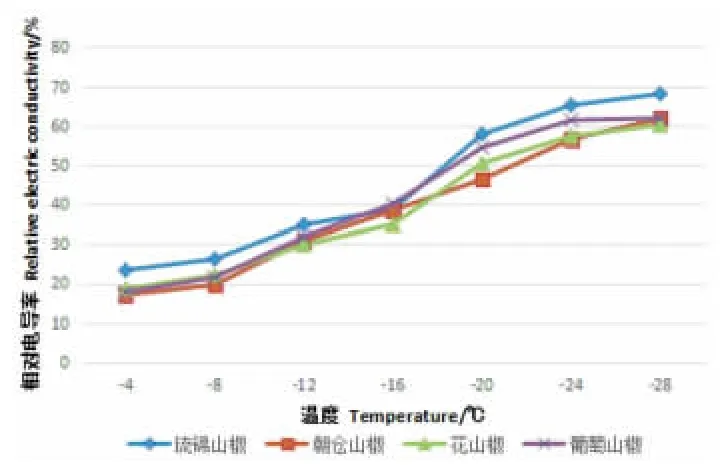
图4 不同低温处理对日本花椒枝条电解质外渗率的影响(单位:%)
3 结论与讨论
(1)正常情况下,细胞内活性氧与防御系统之间保持着平衡。当遭遇低温时,活性氧代谢失调,细胞内的活性氧就会积聚,从而对细胞的正常结构与功能起破坏作用,这时细胞内抗氧化防御系统被诱导和激活,以消除活性氧对细胞的损害。试验结果表明,随着温度的下降,SOD 和POD 和活性呈先下降后升高的变化趋势,这与刘玲等[22]在花椒的研究结果一致,与毕君在山椒的研究结果存在一定差异,这可能是低温处理方法不同所致。SOD 和POD 活性升高,这是4 个山椒枝条对低温胁迫所产生的保护性应激反应。
(2)丙二醛含量与植物抗寒性密切相关,其含量变化可反映植物对低温条件反应的强弱,抗寒性强的品系枝条含量增幅较小,而抗寒性弱的品系枝条含量增幅较大。本试验中,琉锦山椒MDA 含量增幅最大,葡萄山椒的次之,朝仓山椒的再次之,花山椒的最小。说明琉锦山椒的抗寒性最弱,葡萄山椒的抗寒性较弱,朝仓山椒和花山椒的抗寒性较强。
(3)植物遭受低温胁迫,生物膜的结构受到破坏,膜透性增大,从而引起电解质外渗,导致相对电导率的增大。因此,相对电导率大小能反映细胞质膜的损伤程度。细胞质膜透性大小与植物的抗寒性密切相关,同样低温条件下,细胞质膜透性大,电导率高且上升速度快,表明该植物品种抗寒性差,反之,则抗寒能力强。本试验中,当处理温度降低到-16℃,枝条的相对电导率出现了一个快速上升阶段,表明枝条受到了严重伤害,细胞膜透性发生了不可逆的变化。在-16℃降到-20℃时,琉锦山椒电解质外渗最多,脂膜损伤程度最重;葡萄山椒与花山椒电解质外渗较少,朝仓山椒电解质外渗最少。结果表明:朝仓山椒和花山椒抗寒性较强,葡萄山椒次之,琉锦山椒最弱。测定的4 个无刺花椒与毕君[28]的研究结果存在一定差异,这可能是低温处理方法不同所致。
