添加微生物菌剂对菊芋秸秆发酵的影响
2020-05-21魏晓强钟启文张广楠
魏晓强 钟启文 张广楠

摘要:菊芋是青海高原近年来迅速发展的特色经济作物,其地上部秸秆生物量大,为解决其综合利用问题,针对5种微生物菌剂进行了菊芋秸秆静态好氧发酵添加比较研究。结果表明,菊芋秸秆在好氧发酵过程中,添加微生物菌剂能明显促进其发酵进程,有效提高了堆体温度并延长高温持续时间,加快其物质降解与合成,有助于在短时间内使堆料达到理化性质的稳定并消除其植物毒性;添加微生物菌剂还可以提高腐熟堆料的营养水平,减少氮素损耗,保证腐熟秸秆的肥力。5种菌剂促进发酵的效果存在差异,秸秆降解剂008-J表现最优。本研究明确了微生物菌剂对菊芋秸秆发酵的促进作用,为发酵菌剂的筛选和菊芋秸秆基质化利用提供了参考。
关键词:菊芋秸秆;好氧发酵;微生物菌剂;理化性质
中图分类号: S141.4;S182;S188+.4 文献标志码: A 文章编号:1002-1302(2020)06-0246-05
青海省地处青藏高原,高原型大田作物种植种类繁多,每年产生的作物秸秆数量非常可观。随着省内设施无土栽培面积的逐年增加,草炭、蛭石、珍珠岩等成为主要栽培基质,但是这些单一基质是不可再生资源,大量开采会破坏生态环境,因此寻找廉价、清洁、可再生的栽培基质成为无土栽培发展的重点[1]。如果能利用农业废弃物代替草炭等不可再生基质将会带来巨大的经济、生态效益。
菊芋(Helianthus tuberosus L.)是菊科向日葵属多年生草本植物,能适应多种恶劣环境,并且对土壤环境要求不高,因此在青海省高原地区广泛种植。但是,近年来对菊芋的研究主要集中在对地下块茎的研究,而对其地上部研究[2-6]甚少。菊芋地上部茎叶中营养成分较高,并且生物量占全株的50%左右[7]。因此,能有效、环保地处理菊芋秸秆成为一个难题。从农业资源再利用的角度上考虑,解决该问题的最好方法就是对菊芋秸秆进行好氧发酵用作蔬菜栽培基质。刘明池等对菊芋秸秆发酵后的基质化做了研究[8],季延海等在其基础上研究了菊芋秸秆栽培番茄的不同配比[9]。但是对菊芋秸秆发酵的微生物菌剂筛选却没有提到。
本试验利用不同的微生物菌剂来发酵菊芋秸秆,通过对发酵期间的温度、各时期的理化性质、氮磷钾含量与种子发芽指数等进行测定分析,以期筛选出最适合菊芋秸秆发酵的微生物菌剂。
1 材料与方法
1.1 试验材料与处理
菊芋秸秆采自青海大学农林科学院园艺试验基地,将菊芋秸秆粉碎成1 cm左右的小段。5种菌剂分别为郑州益富源生物科技有限公司生产的秸秆发酵菌种008-FZ(T1)、秸秆降解剂008-G(T2)和秸秆降解剂008-J(T3),杨凌万润农业科技有限公司生产的农业用EM菌源(T4)和有机肥专用菌(T5)。菌剂使用前进行扩繁,物料配比见表1,室温下扩繁5~7 d,每天进行搅拌。试验于2017年12月20日至2018年1月19日期间进行,将体积为1 m3的菊芋秸秆堆放在塑料大棚内,分别用微生物菌剂进行发酵,以不加菌剂为对照,扩繁好的菌剂按4 kg/m3的比例加入堆料中进行发酵,每种菌剂设置3次重复。发酵采用室内静态高温好氧的方式,每5 d翻堆洒水1次,保持水分含量在60%左右。室内平均温度为6.3 ℃。
1.2 测试指标与方法
1.2.1 堆体温度 每天13:00对堆体的温度进行测定,测定时确定堆体的3个不同位置,在3个不同位置的40 cm深处进行温度测定并记录,取平均温度作为当天的堆体温度。
1.2.2 理化性质 参照李谦盛的方法测定腐熟秸秆的理化性质[10]。取已知体积(V=650 mL)的烧杯,称质量(m1);将烘干后的秸秆加入烧杯中,称质量(m2);将装有秸秆的烧杯用2层湿纱布封口,完全浸泡在水中过夜(即水要没过容器的顶部),取出称质量(m3),并将湿纱布称质量(m4)。再次用湿纱布将烧杯封口后倒置,让杯内的水自由沥干直至无水流出,称质量(m5)。按以下公式计算各项指标:
1.2.3 酸碱度(pH值)和电导率(EC) 将堆料烘干后粉碎,将堆料和蒸馏水以1 g ∶ 10 mL的比例混合,置于振荡器中于200 r/min振荡30 min,振荡后离心取上清液待用。用ORION STAR A211 pH仪测定pH值,用FiveEasy电导仪进行EC的测定。
1.2.4 全氮(N)、全磷(P)和全钾(K) 全N、全P、全K含量的测定由青海省农林科学院分析测试中心根据国家标准(GB/T 6435—2014《饲料中水分的测定》、GB/T 18633—2018《饲料中钾的测定 火焰光度法》、GB/T T 6437—2018《饲料中钙的测定》)进行测定。用来反映堆体发酵中全N、全P、全K含量的变化趋势。
1.2.5 种子发芽指数(GI) 取堆料鲜样和蒸馏水用1 g ∶ 10 mL的比例混合,置于振荡器中于 200 r/min 振荡30 min,振荡后浸提、过滤取上清液待用。发芽试验材料选用生菜种子,采用培养皿内纸上发芽的方式,每皿内均匀点播30粒种子,加 5 mL 浸提液置于25 ℃人工气候箱内进行发芽试验,每个处理3次重复,以蒸馏水为对照。取样后将生菜种子置于浸提液中72 h,之后测定种子发芽率及发芽种子根长,计算种子发芽指数[11]。
GI=处理的种子发芽率×种子根长对照的种子发芽率×种子根长×100%。
1.3 数据分析
使用Excel 2007、Origin8和SPSS 16.0等数据分析软件对数据进行分析和图表绘制。
2 结果与分析
2.1 堆体温度
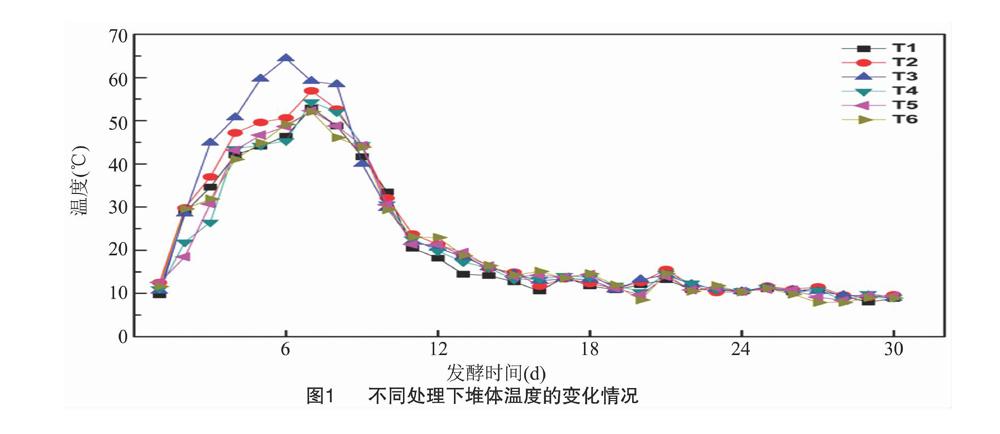
从图1可以看出,添加了发酵菌剂的T1、T2、T3、T4、T5处理与不添加任何发酵菌剂的对照处理(T6)之间温度变化趋势基本相同但也有差别(以下简称T1、T2、T3、T4、T5、T6)。添加T3菌剂的處理温度增长最快,发酵后3 d上升到50.70 ℃,发酵后6 d就达到最高温度(64.37 ℃),在50 ℃以上的高温维持了5 d。说明添加了T3菌剂的处理温度上升最快,最高温度也最高,高温期维持的时间也较长。
2.2 pH值和EC
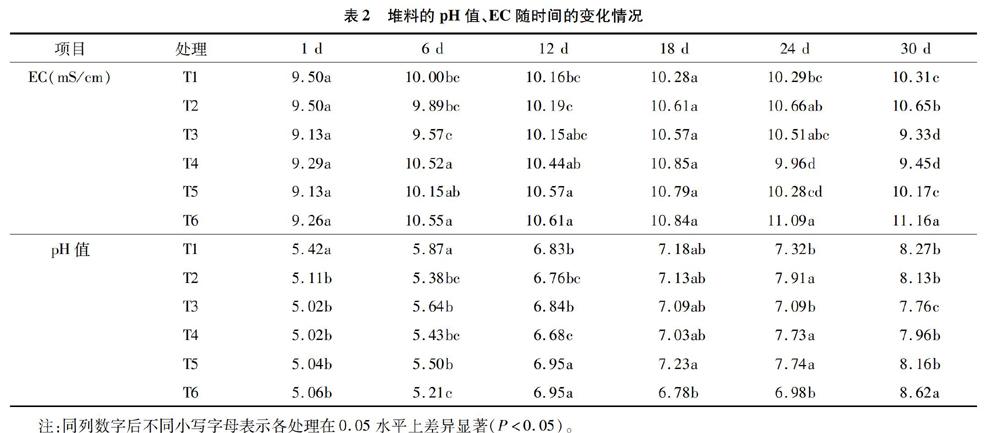
如表2所示,6种处理条件下的堆体在整个过程中 pH 值呈稳步升高的趋势。其中, 添加了微生物菌剂处理的pH值在发酵6 d后均有提高。不加菌剂的处理,堆体pH值在发酵后6 d虽然有所上升,但是较添加菌剂的处理上升不明显。所有处理从发酵后12 d开始pH值上升变缓。发酵后30 d,6个处理的pH值在7.76~8.62之间,T3处理的pH值最小且呈中性。大多数处理发酵结束时的pH值较起始pH值升高3左右。
从各处理EC的变化趋势可以看出,发酵前后菊芋秸秆EC均有所升高但各处理之间变化幅度不同,在堆料发酵的中前期EC呈上升趋势且变化幅度较大,发酵后期T1、T2及T6处理的EC呈缓慢上升的趋势,其余处理堆料的EC逐渐下降。发酵后30 d 6个处理的EC在9.33~11.16 mS/cm之间,T1、T2、T5和T6处理的堆料EC显著高于其他处理。
2.3 理化性质
堆料内大分子有机物的降解造成了堆料容重的变化。如图2所示,6种处理下,T4处理的容重呈先下降后上升又下降的趋势,其余各处理都呈先上升后下降的趋势。而容重的变化与通气孔隙、持水孔隙、气水比的变化息息相关。通气孔隙呈下降趋势而持水孔隙在上升,又因为气水比为通气孔隙与持水孔隙的比值,所以气水比呈下降的趋势。
2.4 全N、全P和全K
N素以NH3的形式挥发是造成堆料内全氮含量变化的主要原因。由图3可知,在好氧发酵的升温及高温期是N素损失的主要时期。高温期后N素通过硝化菌的硝化作用被固定,所以N素的损失逐渐变小,再加上堆体体积和质量的减小,所以堆体的全N含量在后期出现上升的趋势。发酵后 30 d,6个处理的全N含量在11.47~13.68 g/kg之间。6个处理的全氮含量较初始含量均有不同程度的降低,其中T3处理的损失量最小,为0.06 g/kg,这是因为T3处理在升温期迅速升温,减少了升温过程中N的损失。
从堆料的全磷、钾含量的变化趋势可以看出,两者的含量在发酵后均升高,但是添加不同菌剂的处理之间存在差异。各处理在发酵后堆料的全磷含量(以P2O5计)以T3、T4处理的提升辐度较大,分别为0.24、0.26 g/kg。而发酵后全K含量的提升量比全P含量的提升量大,其中以T2、T3处理的提升量较大,分别为4.33、3.57 g/kg,T1、T4处理的提升量次之,T5和T6处理的提高程度差异不大。
2.5 种子发芽指数
如表3所示,发酵至发酵后6 d时,各处理GI均有所下降,说明堆料的毒性增加。但是随着发酵过程中一些小分子物质的降解合成及代谢,堆料中的毒性逐渐消失。发酵后12 d时,T1、T3处理的GI已经大于50%;发酵后18 d时,其余处理的GI均达到50%以上,T3处理的GI接近80%,显著高于同时期其他5个处理。发酵后24 d时,除T5和T6处理外,其余处理的GI均超过80%。发酵后30 d时,T1、T3、T4处理的GI均超过95%,其余处理的GI均超过90%。T5、T6处理的GI在发酵过程中上升的速度较慢,GI在发酵后24 d时才达到70%以上,低于其余处理。发酵30 d后,各处理内种子发芽指数较发酵起始时均显著提高,其中T1、T2、T3、T4处理间无显著差异,均显著高于T5和T6处理。
3 讨论
有研究表明,在堆料进行好氧发酵时添加微生物菌剂能有效增加堆料的微生物数量并缩短发酵过程[12-13]。本研究对堆体发酵过程中的温度变化进行分析发现,添加菌剂能有效缩短堆肥升温的时间且高温期时的平均温度高于不加菌处理,最大高出6.4 ℃,高温持续时间也较对照多1~4 d,最高温则高出0.85~12.32 ℃。这种现象是因为添加了菌剂后,堆体内的初始微生物数量增加,有机物质的分解得到加强,微生物代谢后产生的大量能量使堆体温度迅速升高,缩短了堆体达到稳定期的时间,进而促进了腐熟过程[14]。
栽培基质需要有适宜的理化性质。一般来说,基质容重在0.1~0.8 g/cm3之间,而总孔隙度(通气孔隙与持水孔隙之和)为54%~96%,pH值稍偏酸性为宜[15]。有研究表明,秸秆发酵时不添加碳源会影响基质的稳定性[16]。本研究发现,菊芋秸秆发酵完成时总孔隙度偏大,容重与持水孔隙偏低,堆体pH值在7.76~8.62之间,这可能是因为堆体内没有添加碳源导致的。因此,菊芋秸秆发酵后用作栽培基质时应添加腐熟的羊粪、鸡粪等做为碳源来调节基质的理化性质。
在堆体发酵时,堆料中的含氮物通过微生物的降解形成氨态氮,在高温下以NH3的形式散失,这就造成了堆体总N含量的下降,降低了堆体的肥效[17-19]。在本研究中,添加菌剂的处理较不添加菌剂的处理氮的损失量大大降低,这与前人的试验结果[20]相似,说明在发酵时加入微生物菌剂的确能使全氮的损失减少。
有研究表明,全磷、全钾的含量在发酵前后基本不发生变化[16],但也有人认为磷钾在发酵过程中随着堆体体积的变小其相对含量增加[21]。在本研究中,全磷、全钾的含量都呈上升趋势,发酵后含量明显增加,这可能就是因为其堆体的体积变小导致其相对含量增加,这也能从侧面反映堆料中有机质降解的程度和容重变化情况。
有研究认为,当堆料基本上没有毒性时,GI需要达到50%以上;堆料没有毒性且达到腐熟时,GI需要达到80%以上[22]。而本研究利用生菜种子的发芽试验检验堆料的植物毒性时也得到了相似的结果。在发酵后24 d时,添加了微生物菌剂处理的GI基本达到80%的水平,高于不加菌的处理,且添加不同菌劑的处理之间GI的变化也不相同,T3处理的GI在发酵后12 d就明显高于其他处理。T5处理的GI上升速度总体上低于其余处理,说明T5处理堆料内的微生物代谢不及其他处理。
4 结论
菊芋秸秆在进行好氧发酵时添加微生物菌剂能有效提高堆体的温度,增强菊芋秸秆堆体内物质的合成与降解,缩短植物毒性的解除时间。这说明添加微生物菌剂能有效缩短发酵时间,增加堆体内的微生物数量,促进发酵进程。同时,添加菌剂能减少堆体内全氮的损耗,提高腐熟堆料内的营养水平,保持秸秆的肥力。但不同菌剂之间这种促进效果不一样。本试验结果表明,当添加腐熟菌剂T3时,腐熟效果要优于不加菌和添加T1、T2、T4、T5菌剂的处理。
参考文献:
[1]蒋卫杰,余宏军,刘 伟. 有机生态型无土栽培技术在我国迅猛发展[J]. 中国蔬菜,2000(增刊1):35-39.
[2]Zubr J,Pedersen H S.Characteristics of growth and development of different Jerusalem artichoke cultivars inulin and inulin containing crops[J]. Studies in Plant Science,1993(3):11-19.
[3]邓云波,孙志良,葛 冰,等. 一种新型饲料添加剂——菊糖[J]. 湖南畜牧兽医,2005(6):29-30.
[4]马家津,吕跃钢. 以菊芋为原料利用固定化酶和细胞两步法发酵生产乙醇[J]. 北京工商大学学报(自然科学版),2004,22(6):8-11.
[5]王鹏冬,杨新元,张 捷,等. 菊芋在向日葵育种中的应用[J]. 陕西农业科学,2004(4):38-39.
[6]SánchezJ,Cardona C A.Trends in biotechnological production of fuel ethanol from different feed stocks[J]. Bioresource Technology,2008,99(13):5270-5295.
[7]李 莉,孙雪梅. 青海高原菊芋产业发展探析[J]. 中国种业,2011(9):22-24.
[8]刘明池,季延海,赵孟良,等. 菊芋发酵秸秆复合基质对番茄生长发育的影响[J]. 农学学报,2017,7(1):63-68.
[9]季延海,赵孟良,武占会,等. 番茄栽培基质中菊芋发酵秸秆的适宜配比研究[J]. 园艺学报,2017,44(8):1599-1608.
[10]李谦盛. 芦苇末基质的应用基础研究及园艺基质质量标准的探讨[D]. 南京:南京农业大学,2003.
[11]张亚宁. 堆肥腐熟度快速测定指标和方法的建立[D]. 北京:中国农业大学,2004.
[12]沈根祥,尉 良,钱晓雍,等. 微生物菌剂对农牧业废弃物堆肥快速腐熟的效果及其经济性评价[J]. 农业环境科学学报, 2009,28(5):1048-1052.
[13]王卫平,薛智勇,朱凤香,等. 不同微生物菌剂处理对鸡粪堆肥发酵的影响[J]. 浙江农业学报,2005,17(5):292-295.
[14]席北斗,刘鸿亮,孟 伟,等. 高效复合微生物菌群在垃圾堆肥中的应用[J]. 环境科学,2001,22(5):122~125.
[15]郭世荣. 固体栽培基质研究、开发现状及发展趋势[J]. 农业工程學报,2005,21(14):1-4.
[16]张 晔. 棉秆作为无土栽培基质的前处理技术及其对黄瓜生长的影响[D]. 北京:中国农业科学院,2013.
[17]DeLaune P B,Moore P A,Daniel T C,et al.Effect of chemical and microbial amendments on ammonia volatilization from composting poultry litter[J]. Journal of Environmental Quality,2004,33(2):728-734.
[18]Raviv M,Medina S,Krasnovsky A,et al.Organic matter and nitrogen conservation in manure compost for organic agriculture[J]. Compost Science & Utilization,2004,12(1):6-10.
[19]Raviv M,Medina S,Krasnovsky A,et al.Conserving nitrogen during composting[J]. Biocycle,2002,43(9):48-51.
[20]马丽红,黄懿梅,李学章,等. 两种添加剂对牛粪堆肥中氮转化及相关微生物的影响[J]. 干旱地区农业研究,2010,28(1):76-82.
[21]张 蓓. 碳氮比及腐熟菌剂对玉米秸秆发酵的影响[D]. 兰州:甘肃农业大学,2012.
[22]Riffaldi R,Levi-Minzi R,Pera A,et al.Evaluation of compost maturity by means of chemical and microbial analyses[J]. Waste Management & Research.1996,4(1):387-396.杨 娟,郭青霞. 基于水文模型对比建立SWAT模型数据库——以黄土丘陵沟壑区岔口流域为例[J]. 江苏农业科学,2020,48(6):251-2
