红掌大小孢子发生与雌雄配子体发育
2020-05-20李心杨柳燕顾俊杰
李心 杨柳燕 顾俊杰


摘要 选用石蜡切片法观察了红掌品种Pandola的大小孢子发生及雌雄配子体发育过程。研究结果表明,红掌子房为二心皮、二心室,每个子房有2个直生胚珠,胚珠为双珠被,厚珠心,胚囊发育为廖型胚囊,成熟胚囊为八核七细胞胚囊,包括2个助细胞、1个卵细胞、2个极核、3个反足细胞,其中反足细胞有退化趋势;红掌花药4室,每侧有2个花粉囊,花药壁从外至内由表皮、药室内壁、中层和绒毡层组成,绒毡层为变形绒毡层类型,小孢子减数分裂类型为连续型,成熟花粉为2-细胞型花粉粒;红掌大小孢子与雌雄配子体发育状态可以由花梗与叶柄的长度比、苞片与花梗的夹角和苞片的颜色来辅助判断。
关键词 红掌;孢子体发生;配子体发育;石蜡切片
Abstract Sporogenesis and gametogenesis of Anthurium andraeanum cv.Pandola were investigated by techniques of paraffin sections.The main results are as follows:The ovary of A.andraeanum is bicarpellary and bilocular.Each ovary has two orthotropous ovules,which is bitegmic and crassinucellate.The development of embryo sac is of an Polygonum type.The mature embryo sac is 7celled and 8nucleate type,including two synergids,one egg cell,two polar nucleis and three antipodal cells,and the antipodal cells have a degradation trend.The anthers of A.andraeanum are tetrasporangiate.Each side has two sporangials.The anther wall is composed of epidermis,endothecium,middle layer,tapetum from outside to inside.The tapetum belongs to a deformation type.The cytokinesis in the microsporocyte meiosis is successive,and the mature pollen grains are 2celled.The stage of sporogenesis and gametogenesis of A.andraeanum can be inferred by the length ratio of pedicel to petiole,the angle between bract and pedicel and the color of bract.
Key words Anthurium andraeanum;Sporogenesis;Gametogenesis;Paraffin sections
紅掌(Anthurium andraeanum Linden.),又名花烛、安祖花等,为天南星科花烛属多年生草本花卉,因花型独特,佛焰苞鲜艳亮丽,色彩丰富,花期长,叶型别致,观叶观花俱佳,已成为仅次于热带兰的第二大热带花卉种类,也是我国目前流行的花卉之一[1-3]。我国自20世纪90年代规模化栽培红掌,目前其已成为国内多数地区的重要设施栽培切花和盆栽花卉,并有不断普及的趋势。但我国的主要栽培品种种苗主要从国外引进,自主产权的品种偏少,根据红掌生长开花对环境条件的要求,需要配置较强的设施环境条件,造成国内多数地区设施栽培的加温、降温等成本居高不下。因此,加快培育适宜我国不同气候条件的自主产权品种显得尤为迫切。
红掌的育种进程中,杂交选育发挥了重要作用,目前依然是新品种培育的主要手段。国内的研究主要集中在种质资源[4-6]、栽培管理[7-8]、抗性生理[9-10]和种苗繁殖[11-13]等方面,而杂交选育研究处于起步阶段[14-16],在花发育和生殖生物学等领域的系统研究报道较少。红掌生长到一定阶段后呈现“一叶一花”的连续开花与生长发育模式,花序为肉穗花序,雌蕊发育与成熟期早于雄蕊,有必要对发育进程进行显微观测研究。
该研究采用石蜡切片法,对红掌大小孢子发生和雌雄配子体发育进行显微观测,并对发育进程与形态特征进行对应分析,为红掌杂交育种和胚胎学研究提供重要依据。
1 材料与方法
试验材料为红掌品种Pandola(A.andraeanum cv.Pandola)开花期植株,种植于上海市农业科学院温室大棚,塑料盆栽培,基质为进口泥炭,参照相关技术进行栽培管理。
2018年春季,根据观赏器官苞片的不同发育状态与长度大小,分批采集靠近苞片处的肉穗花序,FAA固定液(90 mL 70%乙醇、5 mL冰醋酸,5 mL福尔马林)固定,常规石蜡切片法制片,切片厚度8~12 μm,爱氏苏木精片染30 min,自来水水洗反蓝,经脱水透明封片后,Nikon光学显微镜下进行大小孢子发生和雌雄配子体发育过程观测,并拍照记录。
2 结果与分析
红掌为肉穗花序,花序上着生小花均为两性花,每朵花具4花被、4雄蕊、2心皮。花序上小花发育进程不一致,底部发育早,顶部发育晚,从下至上依次成熟。形态观测发现红掌柱头分泌粘液的时间早于雄蕊散粉15~20 d。
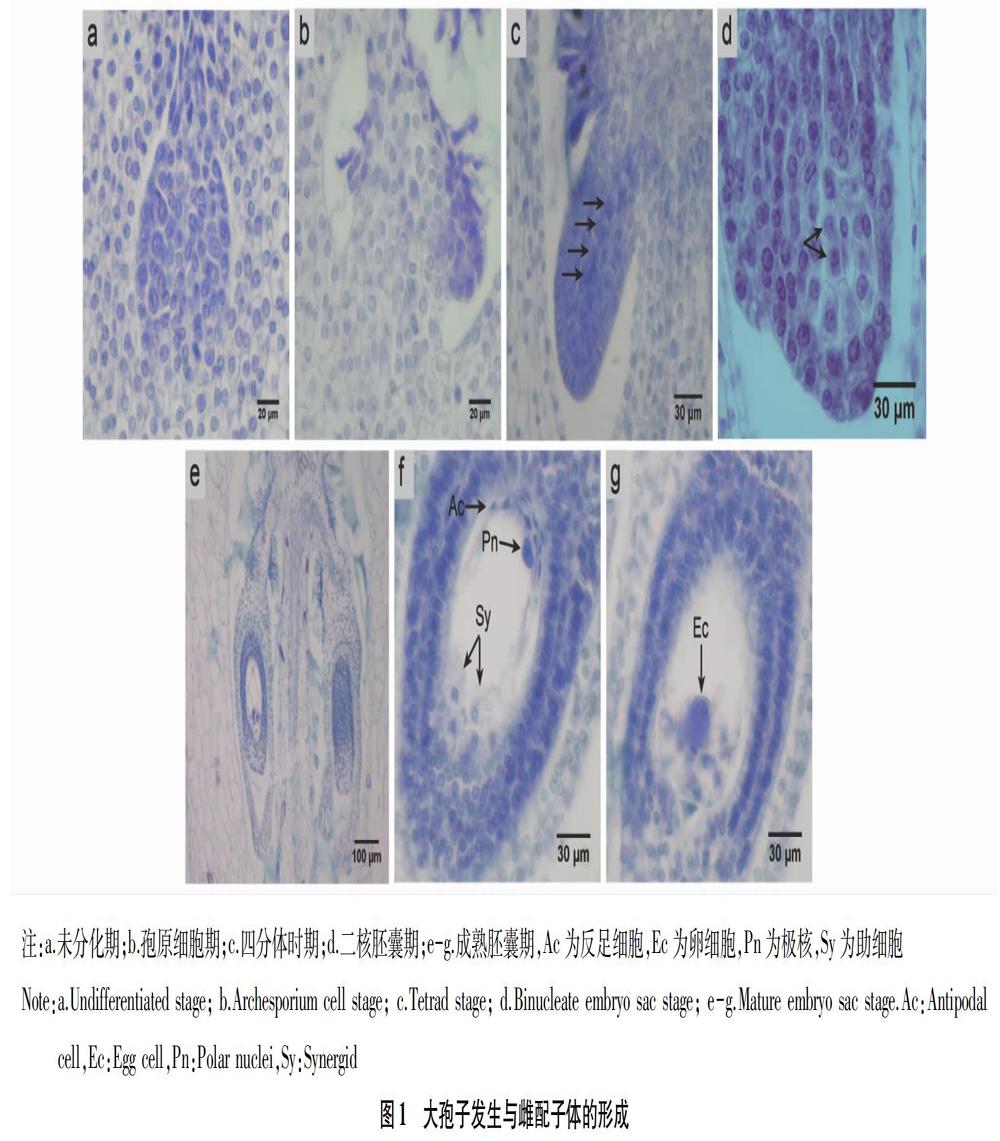
2.1 大孢子发生与雌配子体发育进程 Pandola肉穗花序基部的子房石蜡切片结果表明(图1),红掌子房为二心皮,二心室(图1a);每个子房有2个直生胚珠,胚珠为双珠被,厚珠心(图1e);胚囊发育为廖型胚囊(图1c);成熟胚囊为八核七细胞胚囊(图1f~g),包括2个助细胞、1个卵细胞、2个极核、3个反足细胞。
子房发育早期,胚珠原基产生于心皮的腹缝线处(图1a),细胞大小较为均匀一致,随后表皮内部出现一个体积较大细胞,其液泡较大,核仁明显,即为孢原细胞(图1b),孢原细胞直接发育为大孢子母细胞,并经2次减数分裂产生4个大孢子细胞(图1c),其中靠近珠孔端的3个细胞退化消失,合点端大孢子细胞则继续发育,经3次有丝分裂后形成八核胚囊,第1次有丝分裂形成的2个核分别移至两极,从而建立了极性,当第3次有丝分裂完成后,珠孔端和合点端各有4个核,其中各有1个核移至中央构成2个核的中央细胞,另外珠孔端的三核分别发育成1个卵细胞和2个助细胞,合点端的三核均发育成反足细胞,并在成熟胚囊中有退化趋势。
2.2 小孢子发生与雄配子体发育
Pandola肉穗花序基部的花药石蜡切片结果表明(图2),Pandola红掌花药具有4个花粉囊,中部为药隔,每侧有2个花粉囊(图2b),在成熟后合并为1室,并开裂散出花粉,成熟花粉为2-细胞型花粉粒(图2g)。
雄蕊原基刚形成时,表皮细胞内是大小基本一致的薄壁细胞,并未开始分化(图2a)。之后由于花药4个角隅细胞分裂较快,使花药变成4个瓣裂状,花药中部形成维管束,并发育成药隔。接着表皮内出现了孢原细胞(图2b),孢原细胞经平周与垂周分裂形成初生壁和造孢细胞群。初生壁进一步分裂与分化形成了4层花粉壁结构,由外至内为表皮、药室内壁、中层、绒毡层。造孢细胞则分化形成小孢子母细胞(图2c)。小孢子母细胞的体积较大,细胞质浓,没有明显液泡。
随后,中层细胞解体,绒毡层也发育至高峰后开始解体,并形成原生质体团移动至花药腔中,营养供应小孢子发育,此类型绒毡层被称为变形绒毡层(图2d~e)。小孢子母细胞进入减数分裂期,经第1次减数分裂形成2分体,第2次减数分裂形成四分体,这2次减数分裂为连续型分裂。之后,四分体胼胝质壁逐渐消失,形成只有1个核的单核花粉粒,即小孢子;刚刚游离出的小孢子,细胞质浓,细胞核位于细胞中央,之后不断吸收解体的绒毡层营养,细胞迅速变大,细胞质中形成了中央大液泡,细胞核随之移到细胞一侧,形成单核靠边期花粉粒(图2f)。之后花粉粒内部单核有丝分裂形成生殖核和营养核,之后不断发育成熟,形成2-细胞型花粉粒。
2.3 大小孢子与雌雄配子体发育过程中的红掌苞叶形态变化关联分析
红掌Pandola品种的大小孢子与雌雄配子体的发育是随着观赏器官苞片的成长而进行的(图3、表1),当苞片顶端刚刚伸出叶柄基部时(图3a),苞片与花梗夹角为180°,苞片颜色为嫩绿色,此时大小孢子均处于未分化期,雌雄蕊原基刚刚形成,心室还未成形;当苞片底端刚刚伸出叶柄基部时(图3b),苞片与花梗夹角仍为180°,苞片色彩开始积累,上部呈浅粉色,下部仍呈浅绿色,小孢子处于孢原细胞时期,大孢子处于未分化期;当花梗长度约为叶柄长度1/3时(图3c),苞片与花梗夹角仍为180°,苞片从上至下均已呈现浅粉色,此时小孢子处于小孢子母细胞时期,大孢子处于孢原细胞期;当花梗长度约为叶柄长度2/3时(图3d),苞片与花梗夹角约为160°,苞片呈现粉红色,此时小孢子发育阶段不完全一致,处于减数分裂期或四分体时期,大孢子则基本处于四分体时期;当花梗长度与叶柄长度基本一致时(图3e),苞片与花梗夹角约为135°,苞片色彩进一步加深,此时雄配子体发育至单核靠边期,雌配子体则处于二核胚囊期;当花梗长度超过叶柄长且佛焰苞盛开时(图3f),苞片与花梗夹角约为90°,苞片着色完成,此时雌雄配子体均发育成熟。
3 讨论
胚胎學特征是植物亲缘关系判断、系统进化进程的重要依据[17]。比较红掌与天南星科的魔芋属[18]、芋属[19]、马蹄莲属[20]的胚胎学特征,发现它们具备很多一致的特征:连续型胞质分裂、变形型绒毡层、2-细胞型花粉粒、双珠被、廖型胚囊等,说明这几个属亲缘关系较近。一般认为,双珠被、厚珠心、廖型胚囊、2-细胞型花粉粒为比较原始的特征[21],而直生胚珠相对倒生胚珠则比较进化[22]。红掌既具有相对原始的特征,又具有相对进化的特征,因此认为花烛属在天南星科中属于由原始向进化过渡的类群。
成功的有性生殖过程保证了植物群体的扩大和育种效率的提高,目前研究表明,大小孢子和雌雄配子体发育进程受到阻碍是植物生殖失败的重要原因。该研究发现,红掌雌雄蕊发育不同步,早期雄蕊先于雌蕊发育,但后期胚囊发育进程极快,使得雌性生殖细胞早于雄性生殖细胞成熟。红掌Pandola品种的大小孢子和雌雄配子体发育进程基本正常,虽有少量败育花粉,但因其花粉量大,并不会影响其结实,因而认为红掌自然结实率较低的原因应是雌蕊比雄蕊先成熟且相差时间较长所导致。
4 结论
该研究以红掌Pandola品种为试验材料,通过形态与切片观测进行了红掌大小孢子和雌雄配子体发育进程观测。结果表明,Pandola红掌子房为二心皮,二心室,每个子房有2个直生胚珠,胚珠为双珠被,厚珠心,胚囊发育为廖型胚囊,成熟胚囊为八核七细胞胚囊,包括2个助细胞、1个卵细胞、2个极核、3个反足细胞,其中反足细胞有退化趋势;红掌花药4室,每侧有2个花粉囊,花药壁从外至内由表皮、药室内壁、中层和绒毡层组成,绒毡层为变形绒毡层类型,小孢子减数分裂类型为连续型,成熟花粉为2-细胞型花粉粒;Pandola红掌大小孢子与雌雄配子体发育状态可由花梗与叶柄的长度比、苞片与花梗的夹角和苞片的颜色来辅助判断。该研究结果为红掌的杂交育种和系统进化研究提供了理论基础。
参考文献
[1] DUFOUR L,GURIN V.Growth,developmental features and flower production of Anthurium andreanum Lind.in tropical conditions[J].Scientia horticulturae,2003,98(1):25-35.
[2] 张永红,王莲英.安祖花生长发育特性初探[J].北京林业大学学报,1995,17(2):73-78.
[3] 易双双,杨光穗,尹俊梅,等.红掌主要观赏性状遗传多样性分析[J].热带作物学报,2017,38(12):2206-2214.
[4] GE Y Y,ZHANG F,SHEN X L,et al.Genetic variations within a collection of anthuriums unraveled by morphological traits and AFLP markers[J].Biochemical systematics and ecology,2012,45:34-40.
[5] 户帅雅,李斌奇,陈孝丑,等.红掌遗传多样性及亲缘关系的ISSR分析[J].福建农业学报,2019,34(12):1-8.
[6] 尚伟,江秋萍,陈昌铭,等.红掌种质资源表型性状的遗传多样性研究[J].安徽农业科学,2016,44(33):1-4.
[7] 尚伟,陈昌铭,林发壮,等.红掌主要农艺性状多样性分析[J].中国农学通报,2019,35(6):129-133.
[8] 左利娟,庞小玲,潘庆辉,等.不同品种红掌植株生长动态研究[J].现代农业科技,2019(20):126-127.
[9] 李心,杨柳燕,张栋梁,等.低温胁迫下2个红掌新种质的耐寒性鉴定[J].分子植物育种,2019,17(16):5446-5453.
[10] 王贵,李惠,周红龙,等.红掌种质细菌性枯萎病抗性鉴定分析[J].江苏农业科学,2016,44(9):138-140.
[11] CHEN C,HOU X L,ZHANG H X,et al.Induction of Anthurium andraeanum “Arizona” tetraploid by colchicine in vitro[J].Euphytica,2011,181(2):137-145.
[12] 盛慧,杜伟玲.红掌高效再生体系及遗传转化体系的建立[J].南方农业,2017,11(7):64-66.
[13] 张晓英,谢炳乾,郭伟.红掌的组织培养技术[J].陕西林业科技,2019,47(5):57-58.
[14] 常娟霞,牛俊海,黃少华,等.红掌种子结实性、发育周期及杂交后代发芽力的变异分析[J].基因组学与应用生物学,2014,33(2):392-397.
[15] 陈春,陈孝丑,张毅智,等.红掌杂交育种与无菌播种技术[J].宁德师范学院学报(自然科学版),2017,29(3):287-292.
[16] 陈艳艳,黄少华,冷青云,等.不同红掌杂交组合F1代重要形态性状的遗传分析[J].热带作物学报,2019,40(8):1488-1494.
[17] JOHRI B M,AMBEGAOKAR K B,SRIVASTAVA P S.Comparative embryology of angiosperms[M].Berlin:SpringerVerlag,1992.
[18] 赵蕾,刘佩瑛.魔芋的胚胎学研究[J].西南农业大学学报,1987,9(2):198-203.
[19] 辛红婵,陆绍椿,王福青,等.芋大小孢子发生和雌雄配子体形成的观察[J].莱阳农学院学报,1990,7(2):125-128.
[20] 杨柳燕,王玉兵,张永春,等.彩色马蹄莲大小孢子发生和雌雄配子体发育的研究[J].植物研究,2015,35(4):504-508.
[21] 玛海希瓦里P.被子植物胚胎学引论[M].陈机,译.北京:科学出版社,1966:53-83.
[22] 霍尔M A.植物的结构、功能和适应[M].姚壁君,等译.北京:科学出版社,1987:100-120.
