西花蓟马P450基因在继代适应菜豆植株中的应答反应
2020-05-11郅军锐岳文波
刘 利,郅军锐,岳文波,谢 文,张 涛
(贵州大学昆虫研究所,贵州省山地农业病虫害重点实验室,贵阳 550025)
寄主植物和植食性昆虫在长期的进化过程中形成了一种相互对抗和彼此适应的机制。被昆虫取食后,植物能产生防御反应抵御植食性昆虫进一步取食为害,植食性昆虫也能通过反防御机制适应寄主植物的防御,确保种群的生存和繁衍(彭露等,2010;刘勇等,2011;禹海鑫等,2015)。细胞色素P450是一类广泛存在于生物体内的超家族蛋白,除与害虫的抗药性有关外(Schuler,2011;Mizutani and Masaharu,2012),在外源有毒物质如次生物质的解毒以及寄主植物的适应性中发挥着重要的作用(邱星辉,2014;陈澄宇等,2015;Liuetal.,2015a)。如烟粉虱Bemisiatabaci(谢文,2011)、二斑叶螨Tetranychusurticae(戴宇婷等,2012)、B型烟粉虱和Q型烟粉虱(杨金键等,2017)在取食不同寄主后,体内细胞色素P450活性发生改变迅速适应不同寄主。Peng等(2016)研究发现烟蚜Myzuspersicae的P450基因CYP6CY4 和CYP6CY3 的表达与次生物质烟碱密切相关。Zhu等(2016)发现马铃薯叶甲Leptinotarsadecemlineata取食植物毒素和药剂处理的马铃薯后,P450s基因表达上调,参与植物毒素和药剂的代谢。花椒毒素也可诱导美洲棉铃虫Helicoverpzea体内P450s基因CYP6B8 和CYP321A1以及珀凤蝶PapiliopolyxenesCYP6B1基因的表达(Petersenetal.,2001;Lietal.,2004)。
西花蓟马Frankliniellaoccidentalis是世界性的重要入侵害虫(Kirk and Terry,2003;Reitzetal.,2011),除直接取食和产卵危害,还能传播多种植物病毒,造成巨大的经济损失(谢永辉等,2013)。西花蓟马于2003年在北京首次报道其为害(张友军等,2003),随后在中国各地扩散(吕要斌等,2011),甚至西藏(王海鸿等,2013)、吉林(刘若思等,2015)和内蒙古中西部地区(高振江等,2017)也有其危害的报道,这可能与其对环境和寄主强大的适应能力有关。因此,探讨西花蓟马对寄主植物的适应性,对揭示其入侵、传播和扩散具有重要意义。
西花蓟马寄主植物广泛,但对不同寄主的适应性不同。菜豆是西花蓟马的嗜食寄主,但菜豆不同部位对西花蓟马的生长发育和繁殖影响不同,西花蓟马对菜豆豆荚的嗜食性高于菜豆植株叶片(郅军锐等,2010)。前人对西花蓟马适应寄主植物机制的报道主要集中在其取食不同寄主(Jensenetal.,2000;刘建业等,2017)或不同诱导的寄主植物后虫体生理生化水平的变化(郅军锐等,2016;蒲恒浒等,2018),而西花蓟马代谢相关基因(如P450s)的应答机制,及逐代适应的调控模式尚不明确。本文在前人研究的基础上,通过实时荧光定量PCR分析了西花蓟马从菜豆豆荚转换到菜豆植株适应不同世代后,其2龄若虫和成虫体内CYP4基因和CYP6相关基因表达量的变化,为深入探讨西花蓟马对寄主植物适应机制提供依据。
1 材料与方法
1.1 材料
供试昆虫:西花蓟马为人工气候箱中长期继代饲养(20代以上)的菜豆豆荚种群。条件为温度25±1℃,相对湿度75%±5%,光周期14 h ∶10 h(L ∶D)。
供试植株:矮生菜豆Phaseolusvulgaris品种:金束鹿泰国地豆王(河北省辛集市盛农种子公司)。于人工气候室内种植菜豆植株,条件为温度25±1℃,相对湿度75%±5%,光周期14 h ∶10 h(L ∶D)。每营养钵(直径10 cm,高9 cm)培育两株清洁苗,待菜豆长至3~4真叶时,取长势一致的菜豆植株作为供试寄主植物。生长期间不施用任何农药。
1.2 方法
1.2.1菜豆植株上不同世代西花蓟马的饲养及2龄若虫和成虫的取样
选取健康、长势一致的菜豆植株置于人工气候室的养虫笼(86 cm×45 cm×41 cm)中,取菜豆豆荚上饲养的羽化后3 d并已交配的西花蓟马雌成虫接到菜豆植株上,24 h后剔除所接所有成虫,待西花蓟马产下的卵发育至2龄若虫阶段,每个养虫笼取200头若虫为一个样,放入1.5 mL离心管中,记为F1代2龄若虫,并立即用液氮冷冻后保存于-80℃冰箱中备用,同时保留一定若虫继续在植株上饲养,至成虫阶段后取成虫150头放入1.5 mL离心管中,记为F1代成虫。
取F1代已交配的雌成虫,依照F1代相同的方法转换到新的菜豆植株上,待发育到2龄若虫和成虫后进行取样,分别记为F2代2龄若虫和F2代成虫。按照以上方法继续取F3代2龄若虫和F3代成虫。每一个养虫笼的蓟马为1个重复,共设置5个生物学重复。以取食豆荚的西花蓟马2龄若虫和成虫为对照,记为CK。
1.2.2西花蓟马总RNA的提取
参照Eastep® Super总RNA提取试剂盒说明书步骤提取西花蓟马2龄若虫和成虫的总RNA,并保存于-80℃冰箱内备用。
1.2.3cDNA第一条链的合成
参照RevertAid First Strand cDNA Synthesis Kit试剂盒说明书步骤合成cDNA第一条链,-20℃冰箱内保存备用。
1.2.4实时荧光定量(Realtime qPCR)分析
根据西花蓟马已克隆的5个CYP4基因(董红刚等,2015)和EST数据库中的4个CYP6基因CYP6-1(GQ290643)、CYP6-2(GQ29064)、CYP6-3(GT298397)和CYP6-4(GT298649)(谢志娟,2013)合成的qPCR特异性引物(见表1),以西花蓟马EF-1作为内参基因(Zhengetal.,2014)进行实时荧光定量PCR(RT-qPCR)。引物由生工生物工程(上海)股份有限公司合成。
反应在CFX96®实时荧光定量PCR仪(Bio-rad公司)上进行。反应体系10 μL:5.0 μL FastStart Essential DNA Green Master,正反向引物各0.5 μL,cDNA 1 μL,DEPC水3 μL。程序为:95℃预变性10 min,95℃变性30 s、60℃退火和延伸30 s,进行40个循环。每次生物重复进行3次技术重复。反应完成收集Ct值,分析溶解曲线。
1.3 数据分析
结果采用2-ΔΔCt法(Livak and Schmittgen,2001)进行基因相对表达量的计算。用Excel 2016和SPSS 19.0软件对实验数据进行统计和分析,不同世代间差异性分析用Duncan氏多重比较,2龄若虫和成虫间的差异采用独立样本t检验。

表1 RT-qPCR所用特异引物Table 1 Primers used in RT-qPCR
2 结果与分析
2.1 西花蓟马转换到菜豆植株后体内CYP4基因差异表达分析
对于CYP4基因,西花蓟马从菜豆豆荚转换到菜豆植株后,2龄若虫体内CYP4-1和CYP4-2的表达量变化相似,均是在F1代显著升高,分别为对照的4.37倍和2.31倍,而在F2和F3代其表达量均下降但仍高于对照,且F2和F3代间无显著差异。2龄若虫CYP4-3的表达量只有F1代显著升高,其余世代与对照则无显著差异。CYP4-4表达量仅在F3代显著高于对照,为对照的13.61倍,其余世代与对照无显著差异;CYP4-5表达量的变化呈现先下降后上升的变化,在F2代达到最低值,仅为对照的44.62%,且F1与F3代表达量相当,显著低于对照,分别只有对照的73.04%和68.18%。
成虫体内CYP4-1、CYP4-2和CYP4-3在F1代的表达量与对照相比均无显著差异;而CYP4-1和CYP4-2的表达量在F2代显著下降到最低,到F3代有所上升但仍显著低于对照;CYP4-3的表达量则随世代的增加逐渐降低;CYP4-4和CYP4-5的表达量在F1代显著升高,而F2和F3代与F1代相比均显著降低,且两世代间差异不显著,其中CYP4-4下降到对照的表达水平,而CYP4-5的表达量则显著低于对照。
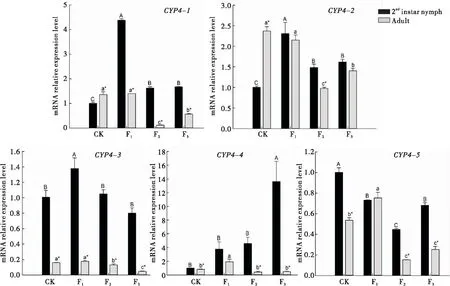
图1 西花蓟马CYP4基因的相对表达量Fig.1 Relative expression levels of CYP4 gene in Frankliniella occidentalis注:图中数据为平均值±标准误,大写字母表示西花蓟马2龄若虫体内CYP4基因表达量在不同世代间显著差异;小写字母表示西花蓟马成虫体内CYP4基因表达量在不同世代间的差异显著性(Duncan氏多重比较,P<0.05)。*表示同一世代西花蓟马体内CYP4基因的表达量在2龄若虫和成虫之间存在显著差异(t 检验,P<0.05)。下图同。Note:Data in the figure are expressed as mean±SE.Capital letters indicate significant differences at the level of 0.05 of gene expression in 2nd instar nymphs among different generations,while lowercase letters indicate significant differences at the level of 0.05 of gene expression in adults among different generations (Duncan’s multiple range test).* indicated that there were significant differences at the level of 0.05 between the 2nd instar nymphs and adults at the same generation (t test).The same for the following figures.
同一个基因的表达量在西花蓟马成虫与2龄若虫间不同。取食菜豆豆荚的西花蓟马成虫体内CYP4-1和CYP4-2的表达量显著高于2龄若虫,而CYP4-3、CYP4-4和CYP4-5的表达量相反,均显著低于2龄若虫的。转换到菜豆植株后只有CYP4-2在F1和F3代,以及CYP4-4和CYP4-5在F1代的西花蓟马2龄若虫和成虫之间无显著差异,其余CYP4基因在2龄若虫的表达量均显著高于成虫。
2.2 西花蓟马转换到菜豆植株后体内CYP6基因差异表达分析
从豆荚转换到菜豆植株后,西花蓟马2龄若虫体内CYP6基因的表达量在不同世代间的变化不同。CYP6-1的表达量在取食菜豆植株后只有F2代显著降低,而CYP6-2表达量只有在F3代才显著升高;CYP6-3 的表达量在取食菜豆植株后均显著低于对照,且在F2代最低;而CYP6-4的表达量只有在F1代才明显升高。
转换到菜豆植株后,西花蓟马成虫体内CYP6-1、CYP6-2和CYP6-3的表达量变化相似,都是在F1代显著下降,与对照相比分别降低了39.55%、39.54%和26.90%,且3个基因在F2和F3代间的表达量均无明显差异,但均显著低于F1代,CYP6-4的表达量在F1代显著升高,但F2、F3代与对照无显著差异。
西花蓟马从菜豆豆荚转换到菜豆植株后,除CYP6-3的表达量在F1代的2龄若虫和成虫间差异不显著外,在其余情况下均是2龄若虫体内的表达量显著高于成虫。
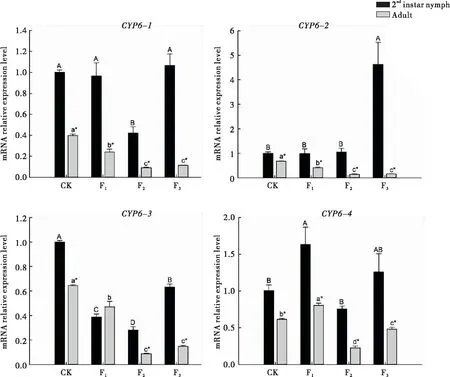
图2 西花蓟马CYP6基因的相对表达量Fig.2 Relative expressionlevels of CYP6 gene in Frankliniella occidentalis
3 结论与讨论
细胞色素P450是一类由CYP基因编码的多基因超家族酶(Werckreichhart and Feyereisen,2000;Feyereisen,2006),对昆虫体内多种内源性和外源性物质的氧化代谢起着重要作用,涉及昆虫的生长发育、营养、对植物毒素的耐受性和对杀虫剂的抗药性等(Scott and Wen,2001;Bonoetal.,2008)。本研究发现西花蓟马从嗜食性很高的菜豆豆荚转换到嗜食性次之的菜豆植株后,2龄若虫体内CYP4-1、CYP4-2、CYP4-3、CYP6-4和成虫体内CYP4-4、CYP4-5、CYP6-4的表达量均在F1代显著上升,以迅速适应新的寄主。这可能是由于西花蓟马转换到菜豆植株后,诱导植物产生有毒次生物质和防御蛋白,对西花蓟马产生不利影响(田甜等,2014;郅军锐等,2016;岳文波等,2018),从而导致其解毒酶基因发生不同的变化。前人也有类似的结果,如周新改(2011)发现烟粉虱转换到新寄主甘蓝后,虫体P450基因表达量上升。槲皮素、呋喃香豆素和花椒毒素等多种植物次生物质也能够诱导昆虫P450不同基因的表达(Lietal.,2004;Maoetal.,2009;陈澄宇等,2015)。彭天飞(2016)发现CYP6CY3 参与代谢次生物质烟碱,其表达量与烟蚜寄主适应性相关。棉铃虫Helicoverpaarmigera在次生物质槲皮素诱导后,CYP337B1 和CYP6B6 表达量显著升高(Liuetal.,2015b)。Yang等(2011)研究发现褐飞虱Nilaparvatalugens取食抗性品种水稻后,CYP6家族基因CYP6CS1和CYP6CW1 被显著诱导。
西花蓟马2龄若虫体内CYP4-1、CYP4-2、CYP4-3、CYP6-4和成虫体内CYP4-4、CYP4-5、CYP6-1、CYP6-2、CYP6-3的表达量均在F2代之后达到稳定,这与烟粉虱寄主转换后体内α-NA羧酸酯酶和谷胱甘肽S-转移酶活性在F2代基本稳定的结果相呼应(周福才等,2010),这可能是由于昆虫转换到新的寄主或食物后,逐渐适应新的取食环境,致使生理调控达到平衡。而桃蚜取食蚕豆后CYP6a13和CYP6a14的表达量平衡点延迟,分别在第6和7代后才达到稳定(朱经云,2017)。P450表达量变化的差异可能与不同基因、昆虫种类和寄主种类有关。CYP4-5、CYP6-1、CYP6-3在2龄若虫期的表达量和CYP4-1、CYP4-2、CYP6-4在成虫期的表达量变化相似,除CYP6-1在F3代恢复到对照水平外,其他基因表达量在F3代时均低于对照,这说明昆虫解毒酶基因在适应取食新的寄主后,不同基因的表达存在特异性,彼此协调以迅速适应新的环境。
本研究结果还显示,西花蓟马转换到菜豆植株除后CYP4-2的表达量在F1和F3代,以及CYP4-4、CYP4-5和CYP6-3的表达量在F1代两个虫态间差异不显著外,其余情况下均是2龄若虫体内的表达量显著高于成虫的,这可能由于2龄若虫取食量较成虫大,或是由于成虫需要节约能量进行后代繁殖所致。前人的研究也表明P450基因在同一昆虫不同虫态体内的表达量不同,周兴隆等(2016)研究发现,二斑叶螨抗性品系中CYP392E10和CYP392A6的表达量在若螨阶段上调,成螨则不显著,而CYP392A16的表达量各个发育阶段都上调,CYP392D8则在除若螨外的其他阶段都上调。烟粉虱噻虫嗪抗性品系中P450基因p_06013和p_00988在4龄若虫过量表达,而p_05916和p_00478表达量很低且在成虫期过量表达,p_00059和p_00428也在雌成虫阶段过量表达(杨妮娜等,2016)。可见环境胁迫能明显影响昆虫不同发育阶段P450基因的表达量,并且因基因、虫态和环境而异。
本研究探讨了P450基因在西花蓟马豆荚种群转换到菜豆植株后逐代适应中的作用,为进一步研究西花蓟马的适应性提供了依据。但寄主植物和植食性昆虫的相互作用十分复杂,不同食物的“口感”、营养成分以及植物的防御反应(植物次生物质的种类和浓度等)等都会对昆虫造成不同影响,昆虫可通过取食行为、生长发育、生理生化及分子防御等以适应不同寄主(彭露等,2015)。因此西花蓟马对寄主的适应机制还有待进一步深入探讨。
