草地贪夜蛾与斜纹夜蛾解毒相关基因的比较分析
2020-05-11朱克森黄钧鸿冯启理
朱克森,黄钧鸿,冯启理,相 辉
(华南师范大学生命科学学院,昆虫科学与技术研究所,广东省昆虫发育生物学与应用技术重点实验室&广州市昆虫发育与应用技术重点实验室,广州 510631)
草地贪夜蛾Spodopterafrugiperda(Smith,1797) 属于鳞翅目Lepidoptera 夜蛾科 Noctuida灰翅夜蛾Spodoptera的一类原产于美洲热带和亚热带地区的杂食性农业害虫(Todd and Poole,1980;王振营,2019),近来入侵我国云南随后扩散全国呈现出爆发状态,对我国农业生产构成了极大的危害(吴秋琳,2019;张磊,2019)。草地贪夜蛾寄主植物高达353种,主要包括禾本科的玉米、小麦、水稻、高粱、甘蔗以及棉花、甜菜、甘蓝、向日葵、香蕉、甜椒、马铃薯、番茄等多种作物(Montezanoetal.,2018)。草地贪夜蛾由于寄主不同也逐渐分化形成了玉米品系和水稻品系2种单倍型,前者主要取食玉米、棉花和高粱,后者主要取食水稻和各种牧草 (Pashleyetal.,1985;Pashleyetal.,1988;Dumas,2015),这两种外部形态基本一致,但在寄主植物、交配行为以及性信息素成分等方面具有明显不同(Pashely,1987;Cruz-Estebanetal.,2018)。
斜纹夜蛾Spodopteralitura(Fabricius)与草地贪夜蛾同为灰翅夜蛾属农业害虫,斜纹夜蛾广泛分布于亚洲、大洋洲、太平洋西岸、中东等许多国家和地区,也是我国重要农业害虫之一,二者具有较高的生态位重叠,且形态相似,但斜纹夜蛾与草地贪夜蛾存在明显的食性差异,斜纹夜蛾主要为害双子叶植物,寄主植物高达389种,包括棉花、烟草、玉米、甘薯、向日葵、高粱、白菜、甘蓝、芥菜、茄子、番茄、辣椒、冬瓜以及藜科等多种作物(秦厚国等,2006;赵胜园,2019)。
昆虫的广泛性取食与其解毒代谢能力息息相关。在长期侵害过程中,寄主植物与植食性昆虫发生协同进化,害虫在广泛性摄取植物能源的同时,体内强大的解毒代谢防御机制也越渐完善。大多数外源有毒物进入生物体内的解毒过程大致可以分为三个阶段,每个阶段需要不同的解毒代谢酶参与,第一阶段参与的基因家族主要是细胞色素P450(cytochrome P450,P450),主要将这些亲脂性或非极性的异生物素进行氧化,为后续的解毒过程提供底物,第二阶段主要是谷胱甘肽转移酶(glutathione s-transferase,GST)、尿苷二磷酸-糖基转移酶以及羧酸酯酶等,GST主要催化谷胱甘肽和有毒物质扼合而生产毒性较低的扼合物,第三阶段是通过ABC转运蛋白(ATP-binding cassette transporter,ABC transporter)家族将毒素排出细胞 (Despresetal.,2007;Lietal.,2007;Dermauw and van Leeuwen,2014)。
目前,对于草地贪夜蛾与斜纹夜蛾的解毒代谢相关基因仍未得到系统研究,本研究通过对这两种夜蛾科害虫的P450、GST、ABC解毒代谢相关蛋白进行整理与鉴定,以家蚕作为鳞翅目模式生物代表,通过构建系统发育树进行全基因组水平的分子生物学进化分析。该项研究为探索外来物种的危害程度及提供新型绿色防治策略具有重大意义。
1 材料与方法
1.1 解毒代谢相关基因的鉴定
从家蚕基因组数据库(silkworm genome database,SilkDB) (http://silkworm.genomics.org.cn/)下载了家蚕的全基因组序列以及已被注释的家蚕14 623条蛋白质序列(BGIBMGA000001-BGNBMGA014623),从NCBI数据库(ftp://ftp.ncbi.nlm.nih.gov/genomes/all/GCF/002/706/865/GCF_002706865.1_ASM270686v1/GCF_00270 6865.1_ASM270686v1_genomic.fna.gz)下载斜纹夜蛾基因组及注释的基因集;从BIPPA数据库(https://bipaa.genouest.org/data/public/sfrudb/)下载草地贪夜蛾的全基因组序列及gff3注释文件信息,通过脚本提取到21 674条蛋白质序列作为基因集;利用已发表的家蚕、斜纹夜蛾、草地贪夜蛾解毒相关蛋白序列如细胞色素P450、ABC转运蛋白、谷胱甘肽转移酶作为参考基因序列,以blastp对家蚕、斜纹夜蛾、草地贪夜蛾的注释基因集中搜索相关蛋白,其中E值设为10-5,所有得到的P450、ABC、GST序列以手动方式进一步在NCBI数据库逐个blastp确定,对于未确定功能的序列,利用SMART结构域预测平台 (smart.embl.de/) 进行检验和筛选。
1.2 解毒代谢相关基因的系统发育分析以及氨基酸变异位点分析
将最终确定的P450、ABC、GST蛋白序列通过MEGA7.0软件中的ClustW进行多序列比对,将比对后的文件转化为nex格式,通过beast 1.10软件 (Suchardetal.,2018) 使用贝叶斯法构建系统发育树,其参数chain length为10 000 000,logEvery为1 000,burnin为0.1,选取部分基因显著扩张的分枝,通过在线软件PolPhen-2(http://genetics.bwh.harvard.edu/pph2/)预测氨基酸变异位点对蛋白质功能影响。
2 结果与分析
2.1 解毒代谢相关基因的注释与鉴定
对家蚕、斜纹夜蛾、草地贪夜蛾基因组的解毒相关基因进行注释,并对P450、GST、ABC基因数目做出了统计(表1)。在草地贪夜蛾中鉴定出213个P450细胞色素基因,58个GST谷胱甘肽转移酶基因,102个ABC转运蛋白基因;在家蚕鉴定出82个P450细胞色素基因,23个谷胱甘肽转移酶基因,64个ABC转运蛋白基因;而在斜纹夜蛾中鉴定出116个P450细胞色素基因,37个谷胱甘肽转移酶基因,99个ABC转运蛋白基因,值的注意的是,斜纹夜蛾ABC转运蛋白有40个未被注释出来,在进一步确认家族分类的情况下,实际上有1个归类于亚家族A,有14个归类于亚家族B,有3个归类于亚家族C,有2个归类于亚家族D,有17个归类于亚家族E,有1个归类于亚家族F,有1个归类于亚家族G,仅有1个未归于某一类亚家族,而这一现象仅出现在斜纹夜蛾中,草地贪夜蛾与家蚕则均被注释出。统计表明草地贪夜蛾细胞色素P450基因和谷胱甘肽转移酶基因GST数量发生了显著扩张现象。

表1 斜纹夜蛾、家蚕与草地贪夜蛾解毒代谢相关基因数目统计Table 1 Statistics on the number of genes related to detoxification metabolism of Spodoptera frugiperda,Spodoptera litura and Bombyx mori

续表1 Continued table 1
注:根据国际通用的P450命名规则,采用CYP前缀表示细胞色素P450,将氨基酸相似性在40%以上的基因划分为同一个家族,将氨基酸相似性在55%以上的基因划分为同一个亚家族,以大写字母表示亚家族,以数字表示一个特定的基因。昆虫P450基因可以分为4个簇(Clan),分别为CYP2 clan、CYP3 clan、CYP4 clan及线粒体(Mitochondrion)clan。CYP2簇包括CYP1、CYP2、CYP15、CYP17、CYP18、CYP21、CYP303-307、CYP342和CYP343;CYP3簇包括CYP3、CYP5、CYP6、CYP9、CYP28、CYP308-310、CYP317、CYP321、CYP324、CYP329、CYP332、CYP336-338和CYP345-348;CYP4簇包括CYP4、CYP311-313、CYP316、CYP318、CYP325、CYP340、CYP341和CYP349-352;线粒体CYP簇包括CYP11、CYP12、CYP24、CYP27、CYP49、CYP301、CYP302、CYP314、CYP315、CYP333、CYP334、CYP339和CYP353(Feyereisenetal.,2012)。
2.1.1细胞色素P450鉴定与分析
草地贪夜蛾P450基因簇的4个簇基因个数分别为18、100、72、23,而家蚕和斜纹夜蛾分别含有6,28,35,11和7,49,49,9个,通过比较发现,草地贪夜蛾CYP3和CYP4簇基因数量明显高于斜纹夜蛾和家蚕,其中主要在CYP3簇里的CYP6、CYP9以及CYP4簇里的CYP4显著发生了基因家族扩张现象。
对草地贪夜蛾、斜纹夜蛾、家蚕P450基因构建进化树,绝大部分分支bootstrap值高于90(图1)。进化树的分析结果表明,根据P450基因序列的结构和同源性,在进化树中可聚类为4个簇,同时在进化树上也表明CYP6、CYP9、CYP4发生了基因家族扩张现象。
选取P450基因家族中的一显著扩张分支,统计发现草地贪夜蛾共出现7处氨基酸错义突变位点,经预测极有可能影响蛋白质的功能(表2)。对于每一处位点,错义突变在草地贪夜蛾中出现的数量与频率均明显要高于斜纹夜蛾与家蚕,以氨基酸位点325为例(见图2),家蚕中该位置的苏氨酸(T)替换为斜纹夜蛾和草地贪夜蛾中的甲硫氨酸(M),暗示T到M的氨基酸突变可能与夜蛾科昆虫的解毒能力有关,而同为夜蛾科的草地贪夜蛾与斜纹夜蛾相比较下,草地贪夜蛾共有 6个该错义突变位点的拷贝,远高于斜纹夜蛾的1个。
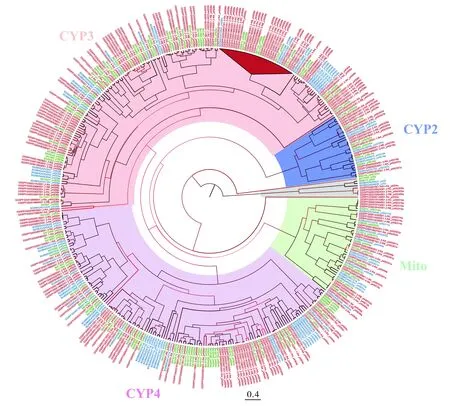
图1 草地贪夜蛾、斜纹夜蛾及家蚕P450的贝叶斯法系统发育树Fig.1 Bayesian phylogenetic tree of P450 in Spodoptera frugiperda,Spodoptera litura and Bombyx mori注:系统发育树分枝颜色的深浅代表分值的高低,三角形代表基因显著扩张部分,阴影区域颜色代表P450基因簇,蓝色代表CYP2,浅橙色代表CYP3,紫红色代表CYP4,浅绿色代表Mito,外层标签颜色代表物种,红色为草地贪夜蛾,绿色为斜纹夜蛾,蓝色为家蚕。Note:Shaded area color represents P450 gene cluster,blue represents CYP2,on behalf of CYP3 shallow orange,purple on behalf of CYP4,light green represents the Mito,outer label color represents species,Red is S.frugiperda,green is S.litura,and blue is B.mori.

图2 草地贪夜蛾、斜纹夜蛾和家蚕中P450_cyp6氨基酸的突变位点分析Fig.2 Amino acid replacements of P450_cyp6 in Spodoptera frugiperda,Spodoptera litura and Bombyx mori注:(A) P450_cyp6蛋白质序列对齐。发生错义突变位点处用红框标识,数字代表突变氨基酸位置。(B) 基于PolyPhen-2软件预测第325位氨基酸残基处的氨基酸替换引起蛋白质功能变化的可能性。家蚕中该位置的苏氨酸(T)替换为斜纹夜蛾和草地贪夜蛾中的甲硫氨酸(M)。PROBABLY DAMAGING,最高级别可能性。Note:(A) Alignment of Protein sequence of P450_cyp6.(B) Possibility of the amino acid replacement at residue 325 to affect protein function predicted by the PolyPhen-2 algorithm.Threonine (T) in B.mori replaced by methionine (M) in S.litura and S.frugiperda .POSSIBLY DAMAGING,the highest degree of possibility to affect protein function.

表2 草地贪夜蛾、斜纹夜蛾、家蚕 P450_cyp6 基因错义替换位点统计Table 2 Non synonymous replacements of P450_cyp6 in Spodoptera frugiperda,Spodoptera litura and Bombyx mori
2.1.2谷胱甘肽转移酶GST鉴定与分析
对草地贪夜蛾、斜纹夜蛾和家蚕GST基因家族进行注释,分别鉴定出58个、37个以及23个GST基因。系统发育树分析结果表明(图3),与斜纹夜蛾相比,草地贪夜蛾GST基因在部分分支中发生了扩张,存在一支中出现5个或6个GST基因成簇聚集,选取GST基因的一成簇聚集扩张分支出现了5处草地贪夜蛾特有的错义突变(表3,图4),这些突变在斜纹夜蛾、家蚕中仅有一个出现,推测这可能与草地贪夜蛾一些独特的解毒能力有关。

图3 草地贪夜蛾、斜纹夜蛾及家蚕 GST 的贝叶斯法系统发育树Fig.3 Bayesian phylogenetic tree of GST in Spodoptera frugiperda,Spodoptera litura and Bombyx mori注:系统发育树分枝颜色的深浅代表分值的高低,三角形代表基因显著扩张部分,外层标签颜色代表物种,红色为草地贪夜蛾,绿色为斜纹夜蛾,蓝色为家蚕。Note:The shade of the branch color of the phylogenetic tree represents the score,the triangle represents the significantly amplified part of the gene,and the outer label color represents the species.Red is S.frugiperda,green is S.litura,and blue is B.mori.

图4 草地贪夜蛾、斜纹夜蛾和家蚕中GST氨基酸的突变位点分析Fig.4 Amino acid replacements of GST in Spodoptera frugiperda,Spodoptera litura and Bombyx mori注:(A) GST蛋白质序列对齐。发生错义突变位点处用红框标识,数字代表突变氨基酸位置。(B) 基于PolyPhen-2软件预测第107位氨基酸残基处的氨基酸替换引起蛋白质功能变化的可能性。家蚕中该位置的酪氨酸(Y)替换为斜纹夜蛾和草地贪夜蛾中的组氨酸(H)。PROBABLY DAMAGING,最高级别可能性的影响。Note:(A) Alignment of Protein sequence of GST.(B) Possibility of the amino acid replacement at residue 107 to affect protein function predicted by the PolyPhen-2 algorithm.Yyrosine (Y) in B.mori replaced by histidine (H) in S.litura and S.frugiperda.POSSIBLY DAMAGING,the highest degree of possibility to affect protein function.

表3 草地贪夜蛾、斜纹夜蛾、家蚕 GST 基因错义替换位点统计Table 3 Non synonymous replacements of GST in Spodoptera frugiperda,Spodoptera litura and Bombyx mori
2.1.3ABC转运蛋白鉴定与分析
对草地贪夜蛾、斜纹夜蛾和家蚕ABC转运蛋白家族进行注释,分别鉴定出102个、99个以及64个ABC基因,根据ATP结合位点的序列相似性,昆虫的ABC转运蛋白被分为8个亚家族(A-H),以亚家族C含量最为丰富,ABCH和ABCG转运蛋白的结构组织相似,亚家族H未被注释出来,有趣的是,斜纹夜蛾未被注释出的40个蛋白有14个归类于ABC转运蛋白亚家族B,另外有17个归类于ABC转运蛋白亚家族E,且都发生了明显的基因扩张现象(图5)。

图5 草地贪夜蛾、斜纹夜蛾及家蚕ABC的贝叶斯法系统发育树Fig.5 Bayesian phylogenetic tree of ABC in Spodoptera frugiperda,Spodoptera litura and Bombyx mori 注:系统发育树分枝颜色的深浅代表分值的高低,三角形代表基因显著扩张部分,外层标签颜色代表物种,红色为草地贪夜蛾,绿色为斜纹夜蛾,蓝色为家蚕。Note:The shade of the branch color of the phylogenetic tree represents the score,the triangle represents the significantly amplified part of the gene,and the outer label color represents the species.Red,S.frugiperda,green,S.litura,and blue,B.mori.
3 结论与讨论
草地贪夜蛾食性杂,寄主范围广,与近缘物种斜纹夜蛾相比,更偏爱禾本科植物如:高粱、小麦、水稻、烟草等,而昆虫的解毒代谢能力与寄主范围以及偏食性有很大的关系 (Ramseyetal.,2010),初步的分析发现,草地贪夜蛾解毒代谢相关基因数目远远大于斜纹夜蛾和家蚕,说明解毒代谢酶的扩张有利于草地贪夜蛾更好的降解不同寄主植物的次生性代谢产物。
细胞色素P450涉及许多昆虫生理功能,包括对信号分子的代谢、宿主植物的适应性及杀虫剂的抗性等,其中最为重要的功能是参与农药、毒素、污染物等解毒的过程(Scottetal.,1998;郭亭亭等,2009),而在草地贪夜蛾的细胞色素P450中,CYP3簇与CYP4簇的基因数目显著扩张,CYP3簇涉及的基因主要参与农药等外源性物质的代谢(Feyereisenetal.,2012),CYP4簇涉及的基因主要的功能则是代谢外源有毒化学物质、参与脂肪酸羟基化、生物合成和代谢等(Feyereisenetal.,2006)。CYP3、CYP4簇中基因数量的显著增多,暗示着草地贪夜蛾的相关解毒能力更为强大,解毒代谢机制更为复杂,能够及时将外界有毒物质进行氧化,使脂溶性物质转变成水溶性物质而被排除体外,更快速高效地降解外源有毒物质。更为有趣的是,我们选取P450 基因中的一显著扩张分支进行序列分析,通过统计发现,在草地贪夜蛾、斜纹夜蛾、家蚕相对保守的氨基酸序列中,草地贪夜蛾共7处位点发生错义突变,经预测这些突变极有可能影响蛋白质的功能。对于每一处位点,错义突变在夜蛾中出现的数量与频率均明显高于家蚕,暗示该突变可能与夜蛾科昆虫的解毒能力有关,而同为夜蛾科的草地贪夜蛾与斜纹夜蛾相比较下,草地贪夜蛾的错义突变位点的拷贝又显著高于斜纹夜蛾,推测草地贪夜蛾通过保留错义突变的解毒酶拷贝并选择将其进行大量扩张,使突变后的解毒酶的相关功能得以加强。
谷胱甘肽转移酶,作为昆虫解毒酶系主要成员之一,主要催化谷胱甘肽和有毒物质扼合而生产毒性较低的扼合物(Sandermannetal.,2012)。GST基因的数量增加与扩张,增强了草地贪夜蛾对植物次生有毒代谢物、农药等的解毒能力,进而导致其在短期内对多种化学农药及Bt产生抗性。对草地贪夜蛾GST基因的聚集扩张分支进行统计,并与斜纹夜蛾、家蚕进行比较,发现了5处草地贪夜蛾特有的错义突变,这些突变在斜纹夜蛾、家蚕中仅有一个突变,推测这可能与草地贪夜蛾一些独特的解毒能力有关。
细胞色素P450和谷胱甘肽转移酶除了作为昆虫解毒酶系的主要成员,也是昆虫气味降解酶主要成员之一,草地贪夜蛾能够识别并定位种类繁多的寄主、更为频繁地交配与产卵,与其嗅觉、味觉受体的扩张相关(黄钧鸿等,2019),但同时草地贪夜蛾也必须具备更强大的气味降解能力,使产生神经冲动后残余的气味分子迅速降解失活,从而保证了昆虫嗅觉系统的高敏感性。因此细胞色素P450和谷胱甘肽转移酶的扩张,可能增加了草地贪夜蛾能够识别的气味分子种类及识别的敏感性,使其大规模迁移与入侵能力在一定程度上优于斜纹夜蛾。
此外,已有研究表明节肢动物的ABC转运蛋白和药物抗性相关(Labbéetal.,2011b),在烟芽夜蛾Heliothisvirescens中发现ABCC2的突变有利于增强Bt毒素抗性(Gahanetal.,2010;Baxteretal.,2011;Atsumietal.,2012)。相对于家蚕,在草地贪夜蛾中ABCC基因家族数量的增加,推测它可能对转基因Bt玉米的抗性与快速适应相关。同时发现,ABCE基因在斜纹夜蛾中也出现了扩张的现象,然而大多数真核生物中只有一个ABCE基因,它在所有生物的进化具有高度的保守性且并没有转运的功能,出现扩张具体的原因有待未来进一步探讨。
总之,草地贪夜蛾解毒代谢相关基因数量的扩张、尤其是某些分支中具有错义突变的蛋白质的基因数量增多,有利于其处理种类更广泛的外源有毒物质、气味物质,并更快速地降解,使其具备更强的解毒能力和气味敏感性,成为其能够在缺乏天敌的情况下快速入侵的重要原因之一,未来可通过进一步对解毒代谢相关蛋白的研究,研发出更具特异性的农药产品,有利于开展对草地贪夜蛾更好的防控措施。
