洞庭湖洲滩土壤种子库对土壤水分变化的响应*
2020-05-08陈明珠雷光春
陈明珠,靳 朝,雷光春,阳 俭,雷 霆*
(1:北京林业大学生态与自然保护学院,北京 100083) (2:河南省出入境检验检疫局,郑州 450003) (3:湖南省西洞庭湖国家级自然保护区管理局,常德 415000)
湿地是单位面积生态服务价值最高的生境类型[1],工业发展、城市化、水利工程等导致湿地生态系统面临多种威胁[2]. 利用种子库技术修复受损湿地是当前湿地研究的热点之一[3]. 研究表明,建设和运用大型水利水电工程,会对下游湿地生态系统的水文节律造成影响[4]. 植物群落的生长发育和繁殖等过程都与水位波动有着密切的关系[5],土壤含水量也是影响湿地植被特征的重要因素[6]. 种子库作为地表植被更新的重要来源,对植物群落受到的破坏能够起到缓冲作用,减少植物种群灭绝的几率[7-8]. 湿地土壤种子库的大小和组成受水文因子、土壤条件[9-10]、湿地类型[11]和地表植被[12]等因素的影响,研究种子库的组成能够了解退化湿地生态系统的物种资源储备[13],还能用于湿地生态系统质量评估、植被演替趋势预测以及植被恢复物种选择等方面的工作[14-15].
土壤种子库受地表植被结构变化及生物学节律影响,其物种组成也会反映地表植物群落的演替趋势[16]. 沼泽湿地及以无性繁殖为主的植被类型中,种子库与地表植被相似性较低[17],潮汐淡水湿地及以一年生为主的植被类型中其相似性较高[18]. 侯志勇等研究了洞庭湖特定区域内虉草、苔草和荻群落地表植被与土壤种子库的相关性[19],Cui等也在太湖研究了4种植物群落地表植被与土壤种子库的相关性[20]. 本次研究立足于洞庭湖当前所面临的生态危机,分别在东洞庭湖、横岭湖、南洞庭湖和西洞庭湖的典型洲滩湿地上采集土壤样品,研究水分胁迫对洲滩湿地种子库萌发特征的影响,分析泥沙洲滩、泥沙-湖草洲滩、湖草洲滩、湖草-南荻洲滩、南荻洲滩5类洲滩地土壤种子库与地表植被的相似性,以期为水文情势变化后洲滩湿地植被的修复策略提供参考.
1 研究区域与研究方法
1.1 研究区域概况
洞庭湖位于长江中游荆江南岸,地处湖南省北部以及湖北省南部,属亚热带湿润季风气候,年平均温度16.4~17.0℃,年平均降水量1200~1550 mm,年平均湿度80%,全年无霜期为260~280 d[21]. 洞庭湖汇集了湘、资、沅、澧四水及长江水资源,再由岳阳城陵矶注入长江,是典型的过水性吞吐型湖泊,多年平均水位24~31 m[22]. 湖区已建立东洞庭湖、西洞庭湖国家级自然保护区,以及横岭湖、南洞庭湖省级自然保护区. 其洲滩湿地沿水岸向内陆依次为泥沙洲滩、湖草洲滩、南荻洲滩及鸡婆柳-旱柳洲滩4种主要类型[10]. 本研究选取的洞庭湖区域主要包括前3种洲滩类型,并在研究过程中将洲滩之间的过渡区域划分为新的类型,即泥沙洲滩、泥沙-湖草洲滩过渡带、湖草洲滩、湖草-南荻洲滩过渡带、南荻洲滩[23].
1.2 植被调查及土壤种子库取样
2013年1月,在洞庭湖4个自然保护区内选取共11处典型洲滩湿地作为样地,其中东洞庭湖5处,横岭湖1处,南洞庭湖3处,西洞庭湖2处(图1,表1).
洞庭湖水位每年自10月逐渐下降,至12-1月达到最低值,来年4月开始水位缓慢上升[24]. 在此水位变化过程中,洲滩湿地面积逐渐增加,在12月到1月之间达到最大值,后缓慢缩小. 我们选择在洲滩面积接近最大值的时期,在每个样地内设置3条由近水裸滩至南荻群落、垂直于水陆交界线的重复样带,样带长度600~1200 m,间距为200~300 m. 采取用空间代替时间的方法设计样品采集方案,每条样带不同部分均包含了洲滩湿地植物从萌发、生长到成熟的各个阶段,采集的混合土壤样品基本包含了各种状态且具有活性的种子.
沿样带布设样方数量为泥沙洲滩1个、泥沙-湖草过渡带1个、湖草洲滩2个、湖草-南荻过渡带1个、南荻洲滩1个,样方间距为100~200 m(图1),样方大小为1 m × 1 m. 调查每个样方内的维管植物种数及个体数,使用WET-2土壤三参数仪测量每个样方土壤含水量. 使用工兵铲铲取每个样方中心位置表层10 cm × 10 cm × 10 cm的土样装入塑封袋密封、标记,带回实验室置于5℃条件下冷藏.

图1 样点分布及样方设置(“█”表示样方) Fig.1 Distribution of sampling sites and quadrat setting (“█” represents quadrat)

表1 样地信息
1.3 种子萌发实验
1.3.1 预处理 采用萌发法,于4月在西洞庭湖国家级自然保护区管理局蒋家嘴分局(28°49′35″N,112°12′11″E)的温室内进行种子库萌发实验,温室内保持通风. 将土样根据每个样带不同洲滩类型混合均匀,过4 mm筛网,除去石块并分拣出土壤中的植物根状茎. 使用四分法将土样平均分成4份,根据不同处理和重复放置于相应萌发盒内. 萌发盒体积为15 cm × 10 cm × 5 cm,底部钻8个孔用于透水. 萌发盒填充物分为3层,最下层铺垫滤纸防止样品和基质掉落,中层为不含有机质的蛭石作为萌发基质,上层放置厚度为2 cm的土壤样品[25]. 已有研究表明,萌发实验中仅有表层1~2 cm的种子会从土壤种子库中萌发[26],而蛭石基质的作用为保持水分和为幼苗根系生长提供空间. 部分植物可依靠根状茎进行营养繁殖,实验中,另将土壤中分拣出的根状茎单独放置于相同规格的萌发盒内进行萌发.
1.3.2 温室实验设置 实验中设置2种处理,每种处理下3个重复,每个重复设置2个平行样本以降低系统误差:① 水淹处理,萌发盒置于大小约60 cm × 45 cm × 45 cm的水箱内,水面没过土样上表面约15 cm;② 湿润处理,样本置于大小约60 cm × 45 cm × 5 cm的托盘内,水面高度约1~3 cm;在温室四角和中间位置放置5个托盘,放置无土壤样品的萌发盒作为空白组,用于排除温室周边植物种子扩散影响. 萌发过程中,湿润处理组和空白组每天加水保持土样湿润,水淹处理组始终保持淹没高度达到15 cm.
1.3.3 数据获取 从有种子萌发开始,每星期记录1次萌发物种的种类及数量. 已鉴定和记录的幼苗当即剔除,避免其产生第2代种子落入种子库;暂时无法鉴定的幼苗将移栽并培养至可鉴定为止. 至不再有新种子萌发,继续观察1个月后结束实验.
1.4 数据处理
1.4.1 种子密度计算 记录各萌发盒内萌发物种数及其幼苗总数,根据土壤取样面积换算为每平方米种子数量作为种子密度指标,本研究中土壤样本采样面积为0.01 m2,每份土样分为2个不同处理和2个平行样本,因此每个样本的种子密度为萌发株数× 400,单位为粒/m2.
1.4.2 Margalef指数 用Margalef指数(R)分析种子库和地表植物物种丰富度:
(1)
式中,S表示植被群落中物种数目,N表示植被群落中全部物种个体总数.
1.4.3 Jaccard指数 采用Jaccard指数(CJ)相似性系数计算种子库与地表植被物种相似程度[11]:
(2)
式中,j为种子库及其对应植物群落共有种,a和b分别为物种数.
1.5 数据分析
利用三因素方差分析(Three-way ANOVA)分析不同洲滩类型、自然保护区和水位处理条件下种子库萌发的密度和物种丰富度的差异,利用单因素方差分析(One-way ANOVA)对5类洲滩地土壤种子库Margalef指数、土壤种子库与地表植被物种组成相似性以及不同水位条件下种子萌发相似性差异进行比较. 若存在显著性差异则采用LSD检验进行多重比较. 采用Pearson相关性分析法分析土壤含水量与种子库大小的相关性,显著度水平为0.05. 采用统计软件SPSS 22.0 (SPSS Inc., Chicago, IL, USA)和Excel 2003进行数据处理,采用Sigma plot 12.5软件 (2011, Systat Software, Inc., USA)进行作图.
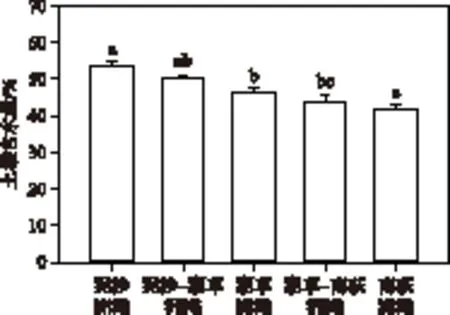
图2 不同类型洲滩土壤含水量(标有不同小写字母表示差异显著,P<0.05)Fig.2 Soil moisture of different kinds of floodplains
2 结果与分析
2.1 各洲滩土壤含水量差异
不同洲滩土壤含水量存在显著差异(P<0.05). 泥沙洲滩距离水面最近,土壤含水量最高,为53.4%;而南荻洲滩远离水面,土壤含水量最低,为41.7%. 沿由水到陆方向,土壤含水量为泥沙洲滩> 湖草洲滩> 南荻洲滩(图2).
2.2 洲滩土壤种子库萌发特征
洞庭湖各类型洲滩土壤种子库密度和物种丰富度均无显著差异(P>0.05). 不同自然保护区和水分处理条件下,土壤种子库萌发的密度和物种丰富度差异显著(P<0.05). 洲滩类型、保护区、水分条件3种因素对土壤种子库萌发的种子库密度和物种丰富度均没有显著交互作用(P>0.05). 洞庭湖4个保护区间洲滩土壤种子库密度和物种丰富度均差异显著(P=0.039、0.018;附录Ⅰ). 其中,东洞庭湖种子库密度最高,南洞庭湖最低(图3a),洲滩种子库萌发的物种丰富度呈现同样的规律(图3b).

图3 洞庭湖4个保护区土壤种子库密度(a)和丰富度(b)(标有不同小写字母表示差异显著,P<0.05)Fig.3 Soil seed bank density (a) and species richness (b) of four nature reserves of Lake Dongting
湿润条件下,萌发的种子库密度显著高于淹水条件下的种子库密度(P<0.01). 湿润组物种丰富度显著高于水淹组物种丰富度(P<0.001). 水淹条件下土壤种子库萌发物种数为11种,其中一、二年生草本5种,多年生草本6种,包括仅在水淹条件下萌发的金鱼藻(Ceratophyllumdemersum)、苦草(Vallisnerianatans)和菹草(Potamogetoncrispus).
2.3 地表植被与土壤种子库的关系
2.3.1 物种丰富度 不同洲滩地表植被物种丰富度差异显著(P<0.05),且由水到陆方向,物种丰富度逐渐增大,泥沙洲滩物种丰富度最小,南荻洲滩物种丰富度最大[23]. 地表植被物种丰富度显著高于土壤种子库(P<0.05),各类型洲滩土壤种子库萌发的物种丰富度没有显著差异(P>0.05;图4). 调查样地内地表植物共36种,湿润条件下种子库萌发物种数32种,其中一、二年生草本23种,多年生草本9种.
2.3.2 物种相似性 不同洲滩类型土壤种子库与地表植被物种组成相似性均小于0.2,且存在显著差异(P<0.05;图5). 其中,泥沙洲滩物种相似性显著低于湖草-南荻过渡带和南荻洲滩地(P<0.05),而后4种洲滩物种相似性差异不显著(P>0.05).

图4 各类型洲滩下地表植物群落和种子库物种丰富度(标有不同小写字母表示差异显著,P<0.05)Fig.4 The species richness of different communities and seed banks of different kinds of floodplains

图5 各类型洲滩下土壤种子库与洲滩地表植被物种组成的Jaccard相似性指数(标有不同小写字母表示差异显著,P<0.05)Fig.5 The values of Jaccard index between seed banks and communities of different kinds of floodplains

图6 地上植被及种子库主要优势种重要值(Pc:蓼子草;Ev:刚毛荸荠;Cs:苔草;Pa:虉草;Ph:芦苇;Ms:南荻;Ch:碎米荠;As:蒌蒿;Lp:陌上菜;Cm:石胡荽;Mj:通泉草)Fig.6 Importance value of the major dominant species in vegetation and seed banks
2.3.3 优势种组成 调查区域内地表植被主要优势种包括蓼子草(Polygonumcriopolitanum)、碎米荠(Cardaminehirsute)等一年生草本植物,以及刚毛荸荠(Eleocharisvalleculosa)、苔草(Carexsp.)、虉草(Phalarisarundinacea)、芦苇(Phragmitescommunis)、南荻(Miscanthussacchariflorus)、蒌蒿(Artemisiaselengensis)等多年生草本植物. 其中泥沙洲滩上长有大量蓼子草,面积最大的湖草洲滩则以苔草为主,而南荻洲滩又以南荻占优. 种子库中主要优势种包括陌上菜(Linderniaprocumbens)、石胡荽(Centipedaminima)、通泉草(Mazusjaponicus)、蓼子草等一年生草本植物以及刚毛荸荠、苔草等多年生草本植物. 其中虉草、芦苇、南荻只在地表植被中存在,而陌上菜、石胡荽、通泉草只在种子库中存在,其余共有种的重要值均存在较大差异(P<0.05;图6).
3 讨论
洞庭湖4个保护区中,东洞庭湖国家级自然保护区种子库密度和物种丰富度明显高于南洞庭湖自然保护区,这可能与保护区湿地保护级别差异及水文和地貌特征的异质性有关. 洲滩湿地土壤含水量存在较为明显的梯度分布,而种子库密度从临近水体到内陆的南荻群落沿高程梯度没有显著差异. 而侯志勇等的研究表明,洞庭湖高水位区的土壤种子库密度大于低水位区,高水位区的一些植物种在洪水来临前完成生活史,种子进入种子库[19]. Greulich等研究了种子库组成与水文和植被类型的关系,发现在较大的洪水梯度上,物种组成显著不同,但种子密度没有差异[27]. 土壤种子库的组成和分布与种子扩散能力[28]、水文和植被、生境条件等因子密切相关[29]. 水文条件影响洲滩植被的分布格局[30],不同植物生产种子的能力不同,种子的性状也有差异,种子性状一定程度决定了种子的扩散能力[31],导致了不同植被类型下种子密度的差异[32]. 种子的扩散能力也会受洪水的季节性和持续时间影响[33-34],洲滩湿地植物的种子主要通过风媒和水媒传播[35],有研究表明,在较小空间尺度内种子扩散能力及进入土壤的数量较为均匀[36]. 此外,湿地中多数种子可以通过洪水再分配,种子漂浮在水面并在水岸边缘聚集,会导致高水位梯度的种子大量积累[37].
不同水分条件处理实验中,湿润条件下萌发物种以陌上菜、石胡荽、通泉草等湿生植物为主,沉水植物因为没有合适的萌发条件而全部缺失,淹水条件萌发物种以沉水植物为主,如金鱼藻、苦草、菹草,湿生植物因水淹条件抑制了萌发,导致它们继续休眠,这一结果验证了淹水状态会影响土壤中种子的萌发能力[26]. 而湖泊的浅水地带和沿岸地带受水位波动的影响最为显著,这些区域即使很小的水位变动都会导致大面积的水体环境与裸露陆地生境之间的转换[38].
本研究中,土壤种子库中萌发的物种以一年生草本植物为主. 由于其存活时间较短,在生活周期内,可以产生大量小而轻的种子,并进入到土壤种子库中[40]. 从低水位的泥沙洲滩区域向高水位的南荻洲滩区域,地表植被物种丰富度逐渐增加,且地表植被与土壤种子库的相似性在长期水淹的泥沙洲滩最低,而在水位波动频繁的泥沙-湖草洲滩相似性最高,这与Schwab等的研究结果相似[41]. Capon等的研究也表明,在洪水更频繁的区域,地表植被与土壤种子库具有更高的相似性[42]. 此外,Touzard等发现,土壤种子库与未受干扰的地表植被相似性较低,地表植被受到干扰后,种子库可以获得一定的萌发空间[43].
洞庭湖各类型洲滩下,土壤种子库与地表植被物种组成相似性总体较低,并且,地表植被物种丰富度普遍高于种子库. 洪水会增加种子库的丰度,但会降低种子库中物种的丰富度和多样性[44]. 已有研究表明,西洞庭湖洲滩湿地土壤种子库与地表植被物种组成的相似性系数介于0.23~0.41之间[45]. 可能由于洞庭湖湿地的苔草、虉草、芦苇、南荻等优势种主要依赖营养繁殖来维持种群规模,种子库中不存在或很少存在这些物种的种子,因此极大降低了地表植被与土壤种子库的相似性[46]. 同时,芦苇和南荻是当地的经济作物,用作造纸原料,在其种子进入土壤种子库前就被收割,也会导致产生的种子部分无法进入土壤种子库[19]. 种子库中陌上菜、石胡荽、通泉草等优势种在地表群落中的重要值均接近0,可能是由于上层苔草或虉草群落盖度过高,阻隔了阳光,而这些物种需要处于阳光下才会萌发[47]. 由于湿地植物对洞庭湖特殊水文环境的适应,洪水后部分主要伴生种如水田碎米荠(Cardaminelyrata)、黄鹌菜(Youngiajaponica)、泥胡菜(Hemisteptalyrata)等物种会快速萌发,仅存在于地表植被,而在土壤种子库中缺少这些物种. 以上这些原因可能共同导致了地表植被的物种丰富度高于土壤种子库,且两者相似性较低.
本研究发现水文条件对洲滩湿地植被的更新潜力起着决定性的作用. 水位升高淹没洲滩会抑制种子的萌发,水位下降洲滩出露则能够促进种子萌发. 这说明在洲滩湿地保护和恢复过程中,水位控制是至关重要的环节,能对洲滩植被的更新潜力和演替模式产生关键性的影响. 同时,地表植被与土壤种子库物种丰富度和相似性的差异表明,仅依靠土壤种子库进行洲滩湿地植被恢复难以达到理想效果,一些湿地中常见的植物如虉草、南荻及部分苔草属植物主要依靠营养繁殖,将会在恢复过程中缺失,需要进行人工引种. 为保证洲滩湿地种子正常萌发,洞庭湖冬季水位需保持在较低水平,湖区水文调控及水利枢纽建设之前应考虑其对洲滩植被的影响.
4 附录
附录Ⅰ见电子版(DOI: 10.18307/2020.0314).
