日本看麦娘对精噁唑禾草灵和甲基二磺隆的抗性机制
2020-04-28毕亚玲戴玲玲李君君
毕亚玲,戴玲玲,谷 刚,李君君
(1.安徽科技学院 农学院,安徽 凤阳 233100; 2.安徽科技学院 资源与环境学院,安徽 凤阳 233100)
日本看麦娘(Alopecurusjaponicus)是冬小麦田主要的恶性杂草之一[1],生产上主要使用精噁唑禾草灵和甲基二磺隆来防治日本看麦娘[2-3]。近年来,由于长期单一地使用精噁唑禾草灵,多个地区小麦田日本看麦娘对其产生较高的抗药性。甲基二磺隆对部分抗乙酰辅酶A羧化酶(ACCase)抑制剂类除草剂的日本看麦娘具有较好的防除效果[4],但近年来已有日本看麦娘等禾本科杂草对其产生抗性的报道[5-7]。杂草的抗药性主要是由靶标和非靶标2个方面的因素产生[8-9]。靶标抗性主要是指植株体内除草剂作用靶标的某个位点发生突变导致的抗药性[10]。王凌越[11]研究发现,6种氨基酸突变会导致日本看麦娘对精噁唑禾草灵产生抗药性,并对相同作用机理的其他ACCase抑制剂产生抗性。非靶标抗性主要是指杂草通过提高体内谷胱甘肽-S-转移酶(GSTs)和细胞色素P450氧化酶等代谢酶的活性,导致到达靶标位点的药剂减少,从而形成抗性[12-15]。韩瑞娟等[16]、裴涛[17]研究发现,GSTs活性增强可导致日本看麦娘对除草剂产生抗性。
目前安徽、江苏、河南等省稻茬麦田日本看麦娘对精噁唑禾草灵和甲基二磺隆的敏感性明显下降,主要表现为田间施用剂量明显上升,但防治效果不好[7,18]。为从源头上了解抗性发生的原因,建立针对性的抗性杂草治理策略,延缓杂草抗药性的发展,本试验拟扩增和比对ACCase和乙酰乳酸合成酶(ALS)基因在日本看麦娘抗性、敏感种群的序列差异,明确靶标抗性机理;通过测定比较日本看麦娘敏感种群与抗性种群中GSTs活性,探究日本看麦娘抗性种群的非靶标抗性机理;研究日本看麦娘抗性种群对多种ACCase和ALS抑制剂的交互抗性,评价其对不同除草剂的敏感性,为抗性日本看麦娘的综合治理提供理论依据。
1 材料与方法
1.1 材料
供试杂草种子:抗性种群AFT采自安徽省六安市寿县安丰塘镇约有12 a精噁唑禾草灵和7 a甲基二磺隆用药历史的冬小麦田。敏感种群AH-7为本实验室已报道的日本看麦娘敏感种群[19],采自安徽省滁州市凤阳县非麦田。试验前期使用整株水平测定法[20]测定抗性种群AFT对精噁唑禾草灵和甲基二磺隆的抗性倍数,分别为44.33和34.43。
试剂:Tris-HCl,上海试剂公司;十二烷基硫酸钠(Sodium dodecyl sulfate,SDS)、乙二胺四乙酸二钠(ethylenediaminetetraacetic acid disodium salt,EDTA-2Na),无锡市亚泰联合化工有限公司;二硫苏糖醇(DTT)、交联聚乙烯吡咯烷酮(PVPP)、聚乙烯比咯烷酮(PVP-40),上海麦克林生化科技有限公司;还原型谷胱甘肽(GSH),湖北弘景化工有限公司;2,4-二硝基氯苯(CDNB),上海浩然生物技术有限公司。
试验所用除草剂:60 g·L-1精噁唑禾草灵(fenoxaprop-P-ethyl)水乳剂,浙江天丰生物科学有限公司;30 g·L-1甲基二磺隆(mesosulfuron-methyl)可分散油悬浮剂,浙江天一生物科技有限公司;15%炔草酯(clodinafop-propargyl)可湿性粉剂,瑞士先正达作物保护有限公司;86.5%烯草酮(clethodim)原药,山东滨农科技有限公司;5%唑啉草酯(pinoxaden)乳油,瑞士先正达作物保护有限公司;70%氟唑磺隆水分散粒剂,浙江天一生物科技有限公司;240 g·L-1甲咪唑烟酸(imazapic)水剂,美丰农化有限公司;100 g·L-1双草醚(bispyribac-sodium)悬浮剂,浙江天丰生物科学有限公司。
仪器:生化培养箱SHP-250,上海坤肯生物化工有限公司;HCL-2000行走式喷雾塔,昆山恒创力科技有限公司;ABI 2720 PCR仪,美国Applied Biosystems公司;H1650R高速冷冻离心机,上海翼悾机电有限公司;JY600E电泳仪,北京君意东方电泳设备有限公司。
1.2 方法
1.2.1 材料培养
选取种子健康状况、成熟度、饱满度等相对一致的日本看麦娘种子,75%乙醇清洗后无菌水漂洗3次,均匀撒在培养皿中无菌水润湿的滤纸上,置于19 ℃生化培养箱中催芽。种子露白后,将其播种到装有壤土的直径为12 cm的塑料花盆中,每盆均匀播种15粒。然后置于温室内培养[20 ℃~25 ℃/10 ℃~15 ℃,12 h/12 h(L/D)],相对湿度65%~75%,自然光照强度。于一叶一心期间苗,每盆留10株株高相近、生长状况良好的幼苗继续培养。
1.2.2 日本看麦娘种群ACCase和ALS基因克隆
日本看麦娘的质体ACCase和ALS基因片段没有内含子,故直接以DNA为模板进行扩增。将日本看麦娘培养至三叶一心期,剪取抗性、敏感种群植株的幼嫩叶片,-80 ℃保存。采用SDS法提取DNA[21]。根据NCBI中GenBank登记的日本看麦娘Alopecurusjaponicus质体型ACCase基因序列(JQ068820.1)和ALS基因序列(AJ437300)分别设计ACCase和ALS基因的扩增引物,进行PCR扩增,扩增引物如表1所示。凝胶电泳鉴定后将PCR产物送往公司测序,对测序结果进行分析。
1.2.3 谷胱甘肽-S-转移酶(GSTs)活性测定
日本看麦娘三叶一心期时,使用HCL-2000行走式喷雾塔分别进行精噁唑禾草灵(有效成分含量62.1 g·hm-2)、甲基二磺隆(有效成分含量9.0 g·hm-2)茎叶喷雾处理,喷雾量450 L·hm-2。药剂用蒸馏水配制成母液,以喷施等量清水作为对照处理,每处理4次重复。施药后继续置于温室内培养,药后第0、1、3、5、7和12天剪取植株地上部分,置于-80 ℃冰箱保存。
表1 扩增日本看麦娘质体型ACCase和ALS基因引物
Table 1 Primers used to amplifyACCaseandALSgene ofA.japonicus

引物Primers序列Sequence(5′-3′)扩增长度Length/bp退火温度Annealingtemperature/℃ACC-FTTTCCCAGCGGCAGACAGAT143755.0ACC-RTCCCTGGAGTCTTGCTTTCAALS-FTCACCAACCACCTTTTCCG165956.0ALS-RCACATTGCACCTTTAGGTCT
GSTs粗酶液提取:取植株组织0.5 g,在液氮中研磨成粉,加入Tris-Hcl缓冲液,参照文献[16]的方法提取GSTs粗酶液。
GSTs活性测定:根据吴进才等[22]的方法,在3 mL 0.1 mol·L-1Tris-Hcl缓冲液中加入0.1 mol粗酶液,25 ℃条件下保温10 min,加入13 mmol·L-1的2,4-二硝基氯苯(CDNB),反应10 min,测量340 nm处的吸光度D值。以未加粗酶液的处理为对照,检测数据为处理的D值减去对照的D值。按照如下公式计算GSTs活性。
GSTs活性(nmol·min-1·mg-1)=D×1000/(c×9.5)。
式中:D为吸光度,c为粗酶液的蛋白质浓度(mg·mL-1)。GSTs相对活性为施药组GSTs活性与同日空白对照组GSTs活性的比值。
1.2.4 AFT种群对不同除草剂的交互抗性测定
测定种群AFT和AH-7对炔草酯、烯草酮、唑啉草酯、甲咪唑烟酸、氟唑磺隆、双草醚的敏感性和抗性倍数。日本看麦娘培养至三叶一心期,使用HCL-2000行走式喷雾塔分别进行除草剂喷雾处理,施药方法参照1.2.3节,除草剂使用剂量见表2,以等量蒸馏水处理为空白对照,各处理3次重复。施药后21 d剪取日本看麦娘植株地上部分,于80 ℃烘箱中烘干至质量不再发生变化,称干质量,计算干质量抑制率。以剂量对数值(x)和干质量抑制率(y)建立回归方程,分别计算各除草剂对抗性种群和敏感种群的有效中量GR50,计算AFT种群对不同除草剂的抗性倍数。
干质量抑制率(%)=[(对照杂草干质量-处理杂草干质量)/对照杂草干质量]×100;

2 结果与分析
2.1 AFT种群中ACCase和ALS的基因突变
表2 除草剂的有效成分用量
Table 2 Active ingredients doses used for determining the sensitivity ofAlopecurusjaponicusto different herbicides g·hm-2
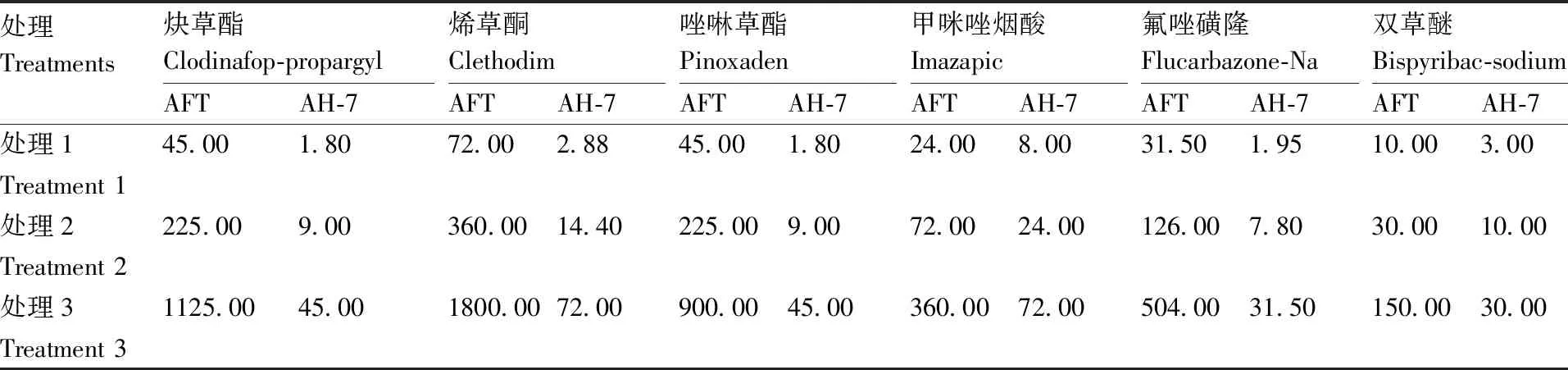
处理Treatments炔草酯Clodinafop-propargylAFTAH-7烯草酮ClethodimAFTAH-7唑啉草酯PinoxadenAFTAH-7甲咪唑烟酸ImazapicAFTAH-7氟唑磺隆Flucarbazone-NaAFTAH-7双草醚Bispyribac-sodiumAFTAH-7处理145.001.8072.002.8845.001.8024.008.0031.501.9510.003.00Treatment 1处理2225.009.00360.0014.40225.009.0072.0024.00126.007.8030.0010.00Treatment 2处理31125.0045.001800.0072.00900.0045.00360.0072.00504.0031.50150.0030.00Treatment 3
在种群AFT和AH-7中扩增ACCase基因的CT区,将测序结果与大穗看麦娘(Alopecurusmyosuroides)的ACCase基因序列进行比对。结果(表3、表4)表明,敏感种群AH-7的ACCase基因CT区2027位氨基酸为色氨酸(TGG),抗性种群AFT的该位置为半胱氨酸(TGC);敏感种群AH-7在ALS基因的574位氨基酸为色氨酸(TGG),抗性种群AFT在该位置为精氨酸(CGG)。
2.2 精噁唑禾草灵、甲基二磺隆对日本看麦娘GSTs活性的影响
图1-A显示,精噁唑禾草灵药剂处理1 d,抗性种群AFT的GSTs活性开始增强,处理3~7 d GSTs活性增加较快,并在第7天达到最高,表明此时抗性植株体内的代谢反应水平被精噁唑禾草灵诱导至最高。处理12 d抗性植株GSTs活性下降,可能是由于植株体内对精噁唑禾草灵的代谢趋于完成,代谢反应水平开始恢复为正常水平。敏感种群AH-7在药后第1~5 d GSTs活性变化平缓,药后5~7 d GSTs活性增强,随后又开始减弱,可能是受到药剂影响或者外界环境胁迫引起的应激反应。
表3 日本看麦娘抗性种群质体型ACCase第2027位的氨基酸突变序列分析
Table 3 Sequence alignment and derived amino acids around position 2027 of the plastidicACCasegene from resistantA.japonicuspopulation
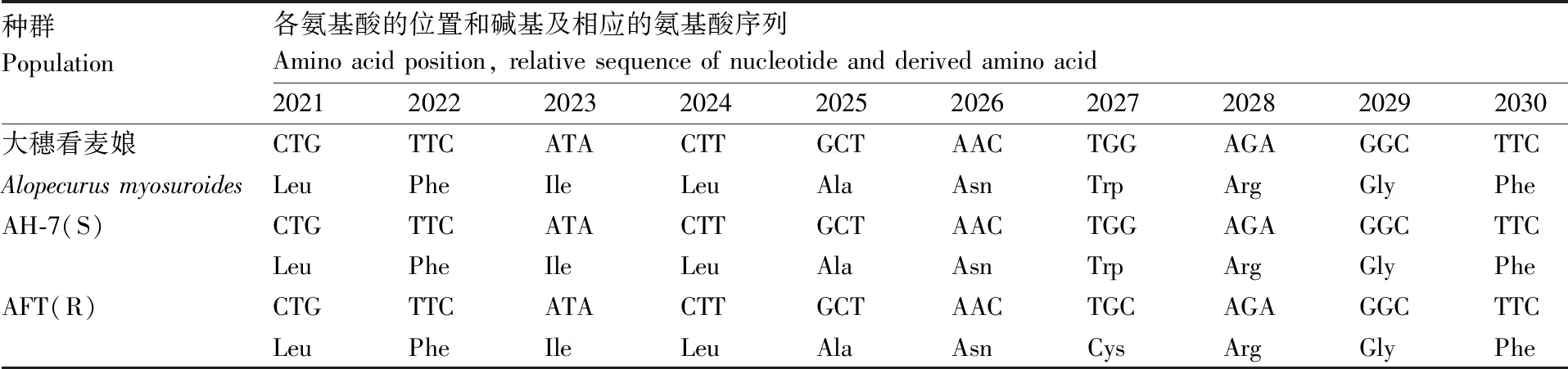
种群Population各氨基酸的位置和碱基及相应的氨基酸序列Amino acid position, relative sequence of nucleotide and derived amino acid2021202220232024202520262027202820292030大穗看麦娘CTGTTCATACTTGCTAACTGGAGAGGCTTCAlopecurus myosuroidesLeuPheIleLeuAlaAsnTrpArgGlyPheAH-7(S)CTGTTCATACTTGCTAACTGGAGAGGCTTCLeuPheIleLeuAlaAsnTrpArgGlyPheAFT(R)CTGTTCATACTTGCTAACTGCAGAGGCTTCLeuPheIleLeuAlaAsnCysArgGlyPhe
氨基酸位数以大穗看麦娘的质体型ACCase全长序列为标准。
Amino acid positions correspond to the full-length plastidicACCaseinA.myosuroides.
表4 日本看麦娘抗性种群ALS第574位氨基酸突变序列分析
Table 4 Sequence alignment and derived amino acids around position 574 of theALSgene from resistantA.japonicuspopulation
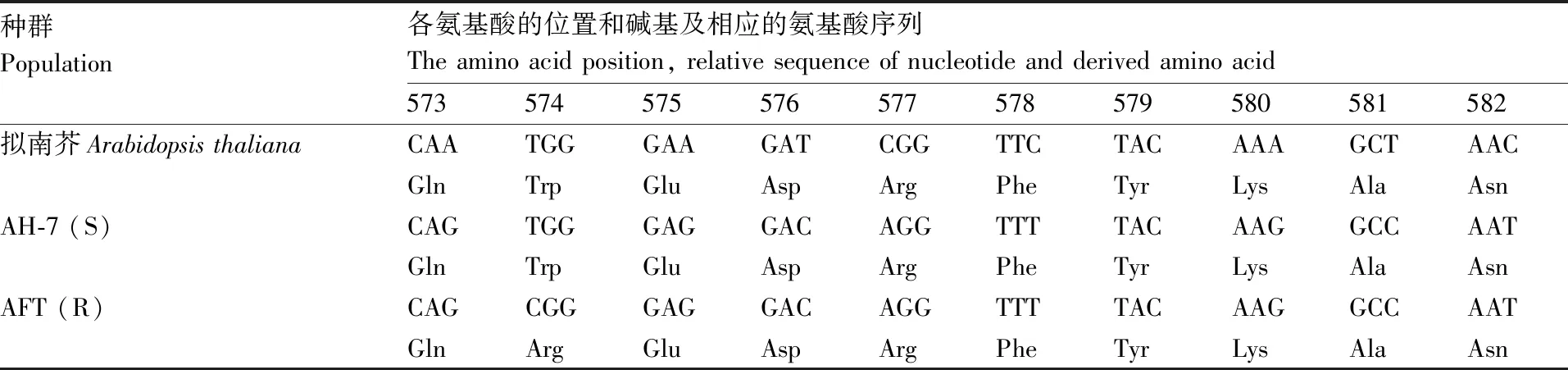
种群Population各氨基酸的位置和碱基及相应的氨基酸序列The amino acid position, relative sequence of nucleotide and derived amino acid573574575576577578579580581582拟南芥Arabidopsis thalianaCAATGGGAAGATCGGTTCTACAAAGCTAACGlnTrpGluAspArgPheTyrLysAlaAsnAH-7 (S)CAGTGGGAGGACAGGTTTTACAAGGCCAATGlnTrpGluAspArgPheTyrLysAlaAsnAFT (R)CAGCGGGAGGACAGGTTTTACAAGGCCAATGlnArgGluAspArgPheTyrLysAlaAsn
氨基酸位数以模式植物拟南芥的ALS全长序列为标准。
Amino acid positions correspond to the full-lengthALSinArabidopsisthaliana.
图1-B显示:甲基二磺隆药剂处理后1~3 d抗性种群AFT的GSTs活性缓慢增强,说明抗性植株体内代谢反应水平开始被诱导升高;药后3~7 d GSTs活性增加较快,并在第7 d达到最大,表明在此时植株体内的代谢反应水平被甲基二磺隆诱导至最高;药后7~12 d,抗性种群AFT的GSTs活性开始减弱,可能是由于植株体内对甲基二磺隆的代谢趋于完成,代谢反应水平恢复至正常水平。甲基二磺隆药剂处理后1~3 d敏感种群AH-7的GSTs活性变化缓慢,药后3 d GSTs活性开始减弱,随后又有增强,在第7天达到最强,12 d又下降。这可能是由于药剂或者外界环境胁迫引起的应激反应,导致GSTs活性出现波动,具体原因还需要进一步的研究。
综合来看,经精噁唑禾草灵和甲基二磺隆药剂处理后,日本看麦娘抗性种群AFT的GSTs活性高于敏感种群AH-7,说明与敏感种群相比,抗性种群体的GSTs对精噁唑禾草灵和甲基二磺隆的代谢能力增强。
2.3 AFT种群对不同除草剂的交互抗性
表5结果显示,AFT种群对炔草酯、烯草酮和氟唑磺隆的抗性倍数分别为27.90、34.43、10.30,表现出高水平的抗性;对唑啉草酯、甲咪唑烟酸、双草醚的抗性倍数分别为5.49、6.42、5.01,表现为中等水平抗性。

同一种群无相同小写字母表示差异显著(P<0.05)。 Data marked without the same lowercase letter of the same populations indicated significant differences at P<0.05.图1 精噁唑禾草灵(A)和甲基二磺隆(B)对日本看麦娘种群GSTs相对活性的影响Fig.1 Effect of fenoxaprop-P-ethyl (A) and mesosulfuron-methyl (B) on relative GSTs activity of Alopecurus japonicas
表5 AFT种群对不同除草剂的交互抗性
Table 5 Cross-resistance in resistant and susceptibleAlopecurusjaponicuspopulations to different herbicides

药剂Herbicide种群Population回归方程Regression equationr有效中量GR5095%的置信区间95% confidence limit抗性倍数Resistance ratio炔草酯AFTy=3.2273+0.7097x0.9674310.28291.97~328.5927.90clodinafop-propargylAH-7y=3.6916+1.2568x0.990011.1210.76~11.481.00烯草酮AFTy=1.8733+1.1165x0.9993637.30595.12~679.4834.43clethodimAH-7y=2.9131+1.6581x0.997918.5117.19~19.831.00唑啉草酯AFTy=2.2586+1.5057x0.994966.4860.11~72.855.49pinoxadenAH-7y=3.4952+1.3662x0.959612.1111.80~12.421.00甲咪唑烟酸AFTy=1.6393+1.4826x0.9739184.06173.14~194.986.42imazapicAH-7y=2.3813+1.8020x0.974928.6526.24~31.061.00氟唑磺隆AFTy=1.8331+1.5898x0.986198.1788.47~107.8710.30flucarbazone-NaAH-7y=3.5739+1.4546x0.98519.538.56~10.501.00双草醚AFTy=1.8561+1.5380x0.9980110.69100.58~120.805.01bispyribac-sodiumAH-7y=3.4924+1.1222x0.956522.1020.48~23.721.00
3 讨论
目前研究表明,杂草的抗药性主要与靶标酶基因突变有关。Guo等[23]报道,在多抗性看麦娘中同时检测到了ACCase基因Ile-1781-Leu和ALS基因Trp-574-Leu 2种突变。本研究使用的抗性日本看麦娘种群AFT采自安徽省六安市寿县安丰塘镇,其对精噁唑禾草灵、甲基二磺隆的抗性均较高,属于多抗性种群。经试验测定,多抗性日本看麦娘种群AFT的靶标酶基因中同时发现ACCase基因CT区基因和ALS基因2种突变类型,这可能是导致种群AFT对二者产生抗性的重要原因。前人研究表明,氨基酸位点突变可能会引起杂草对同一种作用机制的不同除草剂产生交互抗性[24-25]。本试验中抗性种群AFT对其他除草剂产生了不同水平的交互抗性,对炔草酯、烯草酮、氟唑磺隆均产生了较高水平的抗性,对唑啉草酯、甲咪唑烟酸、双草醚产生了中等水平抗性。
非靶标抗性也是导致杂草产生抗性的重要原因[26]。有报道证实了部分杂草的抗药性源自植株内细胞色素P450氧化酶系和GSTs活性增强,从而导致对除草剂的代谢增强[17,27-28]。精噁唑禾草灵和甲基二磺隆处理前,AH-7和AFT种群的GSTs活性差异不明显;药后7 d,抗性种群AFT的GSTs活性显著强于敏感种群AH-7,说明经除草剂诱导后抗性日本看麦娘种群基于GSTs代谢酶介导的代谢反应水平升高,加快了杂草植株对除草剂的代谢解毒反应,推测抗性种群AFT的多抗性可能与植株体内GSTs代谢酶介导的代谢反应机制有关。本试验只测定了GSTs活性,对于非靶标抗性方面还需要进行更深入的研究,比如测定P450氧化酶活性对种群AFT敏感性的影响等。
日本看麦娘种群抗药性的发生除了与自身的生物学特性有关外,与发生地不合理的用药习惯和种植制度也有很大关系。日本看麦娘种群的靶标抗性和非靶标抗性,以及对不同种类除草剂交互抗性的研究,对生产实践中选用不同作用机理的药剂,减小杂草抗药性的发生具有指导意义。农业管理部门应加强对种植户的农技培训,科学合理使用农药,避免出现单一药剂连续使用的情况,同时需要加强对不同作用机理药剂的开发与推广。
