水稻抗稻瘟病基因Pi-ta、Pi-b、Pigm和Pi54在骨干亲本中的分布以及对穗颈瘟抗性的作用
2020-04-22宛柏杰赵绍路朱静雯刘艳艳张桂云朱国永王爱民唐红生孙明法严国红
宛柏杰,刘 凯,赵绍路,朱静雯,刘艳艳,张桂云,朱国永,王爱民,唐红生,孙明法,严国红
(江苏沿海地区农业科学研究所,江苏 盐城 224000)
【研究意义】稻瘟病是由稻瘟病菌(Magnaportheoryzae)引起的水稻重要病害之一,对水稻产量和品质有重要影响,严重时可造成减产40 %~50 %,甚至颗粒无收[1]。选育和利用抗病品种,是防治稻瘟病最经济有效的方法之一,但由于稻瘟病菌存在多个生理小种以及生理小种不断的分化,抗谱较窄的品种往往在推广几年后抗病性就可能出现降低[2-3]。育种实践证明,将具有不同抗谱的稻瘟病抗性基因聚合到同一个品种中,可以有效提高品种抗稻瘟病的持久性[4]。在传统育种中,水稻稻瘟病抗性育种主要依赖于接种鉴定和表型选择,而且需要育种者具有几年甚至十几年的育种经验,鉴定结果往往容易造成误差,甚至抗性基因的丢失,选择效率低,选育周期长[5]。近年来,随着分子生物技术的发展,水稻抗性育种已经进入生物技术与常规技术相结合的阶段,以分子标记辅助选择、多基因聚合等分子育种技术已经成为抗性育种的主要方向[6]。【前人研究进展】目前研究发现,在不同抗病品种中,已经鉴定出80多个稻瘟病抗性基因,其中30多个抗稻瘟病基因已经被克隆[7]。这些基因成簇地分布于除第3 染色体外的其他染色体上[8-10]。随着稻瘟病抗性基因Pi-b第一个被成功克隆,越来越多新的稻瘟病抗性基因通过图位克隆、等位基因挖掘、无毒基因与抗性基因互作等策略得到分离。Pi-b基因位于水稻第2染色体长臂末端附近的区域,编码由1251个氨基酸残基组成的含1个核苷酸结合位点和富含亮氨酸重复的蛋白质[11-12]。Pi-ta基因位于水稻第12 染色体靠近着丝点附近的区域,包含2个外显子和1个1463 bp的内含子,编码1个长度为 928个氨基酸的细胞质膜受体蛋白[13]。Bryan等研究发现Pi-ta基因的编码产物能与稻瘟病菌的无毒基因表达产物相互作用引发抗病反应[14]。Pi54定位于水稻第11染色体上,对稻瘟病菌具有广谱抗性[15]。Sharma等克隆了Pi54基因,在抗病品种Tetep中,Pi54仅包含1个外显子,编码 1个由330氨基酸组成的NBS-LRR类抗病蛋白[16]。Pigm抗性基因来源于我国地方品种谷梅4号,由1对显性基因控制,该基因抗谱广,对9个来自中国不同品系的生理小种均表现出良好抗性[17]。宋兆强等对60 份品系进行了基因检测,解析了稻瘟病抗病基因Pi-ta、Pi-b、Pi54和Pi-km在稻瘟病抗性育种中的作用[18]。大量抗稻瘟病基因的定位与克隆,对通过分子标记辅助选择培育具有持久抗性的水稻品种起到了极大地推动作用。【本研究切入点】本研究利用水稻稻瘟病抗病基因Pi-ta、Pi-b、Pigm和Pi54的分子标记对80份通过传统育种方法选育出来的抗病材料进行基因检测,并结合连续两年人工穗颈瘟接种鉴定结果,对抗病基因进行穗颈瘟抗性评价。【拟解决的关键问题】明确不同抗病基因型对稻瘟病菌的抗病性,有针对性的进行抗性基因的聚合,为筛选广谱抗稻瘟病水稻品种奠定基础。
1 材料与方法
1.1 供试材料
供试水稻材料为江苏沿海地区农业科学研究所经过多年稻瘟病菌圃鉴定筛选出来的抗病育种骨干粳稻亲本80份,依次编号为GP1~GP80。
1.2 稻瘟病抗性基因的标记检测
DNA提取与稻瘟病基因功能标记的检测参考范学军等的方法[19]。抗病基因Pi-ta、Pi-b、Pigm和Pi54引物设计参照陈锋等和吕学莲等[20-21](表1),利用这些功能和连锁标记检测试验材料。利用Pi-ta引物检测到1024 bp片段,并且NPi-ta引物扩增不出目的片段,说明材料含有Pi-ta基因;利用Pi-b引物检测到365 bp片段,并且NPi-b引物扩增不出目的片段,说明材料含有Pi-b基因。而Pi-b引物检测不出目的片段,但NPi-b引物扩增803 bp片段,说明该材料携带有感病基因;抗病基因Pigm引物可以扩增出856 bp大小的目的片段,说明材料中含有抗病基因Pigm,没有扩增出目的基因片段,说明材料不含有Pigm基因;抗病基因Pi54标记为共显性标记,扩增片段216 bp(抗)/359 bp(感)。
1.3 稻瘟病抗性鉴定与评价
1.3.1 抗性鉴定 试验在本所南洋试验基地进行,5月8日播种各试验材料,30 d后移栽,单本载插,每区3行,每行8株,株行距13.3 cm×25.0 cm,整个生育期内不防治病害,虫害防治和肥水管理参考大田生产。
抗性鉴定菌株为江苏省农业科学院植物保护研究所提供,包括近年来江苏省内稻瘟病菌优势小种以及各群小种的代表菌株,2016 年接种鉴定的稻瘟病菌代表菌株: ZB7、ZC11、ZD5、ZE3、ZF1 和ZG1;2017年接种鉴定的稻瘟病菌代表菌株: ZB3、ZC11、ZD7、ZE3、ZF1 和ZG1。将稻瘟病菌株混合,在水稻孕穗至破口期,参照于苗苗等人工注射接种的方法[17],进行水稻穗颈瘟的抗性鉴定,水稻成熟后进行水稻穗颈瘟的抗性调查。
1.3.2 抗性评价 齐穗后30 d调查每个株系的穗发病情况,调查与评价方法参照DB32/T1123-2007《江苏省稻瘟病鉴定方法与抗性评价技术规程》划分抗感类型和病级(表2)。
2 结果与分析
2.1 供试材料抗性基因分析
利用水稻稻瘟病抗性基因Pi-ta、Pi-b、Pigm和Pi54的功能标记对80份育种材料进行检测(图1,表3),结果显示:含有1个抗性基因的材料有12份,占总材料的15 %;含有2个抗性基因的材料有40份,占总材料的50 %;含有3个抗性基因的材料有21份,占总材料的26.25 %;含有4个抗性基因的材料有1份,占总材料的1.25 %;4个抗性基因均没有的材料有6份,占总材料的7.5 %。
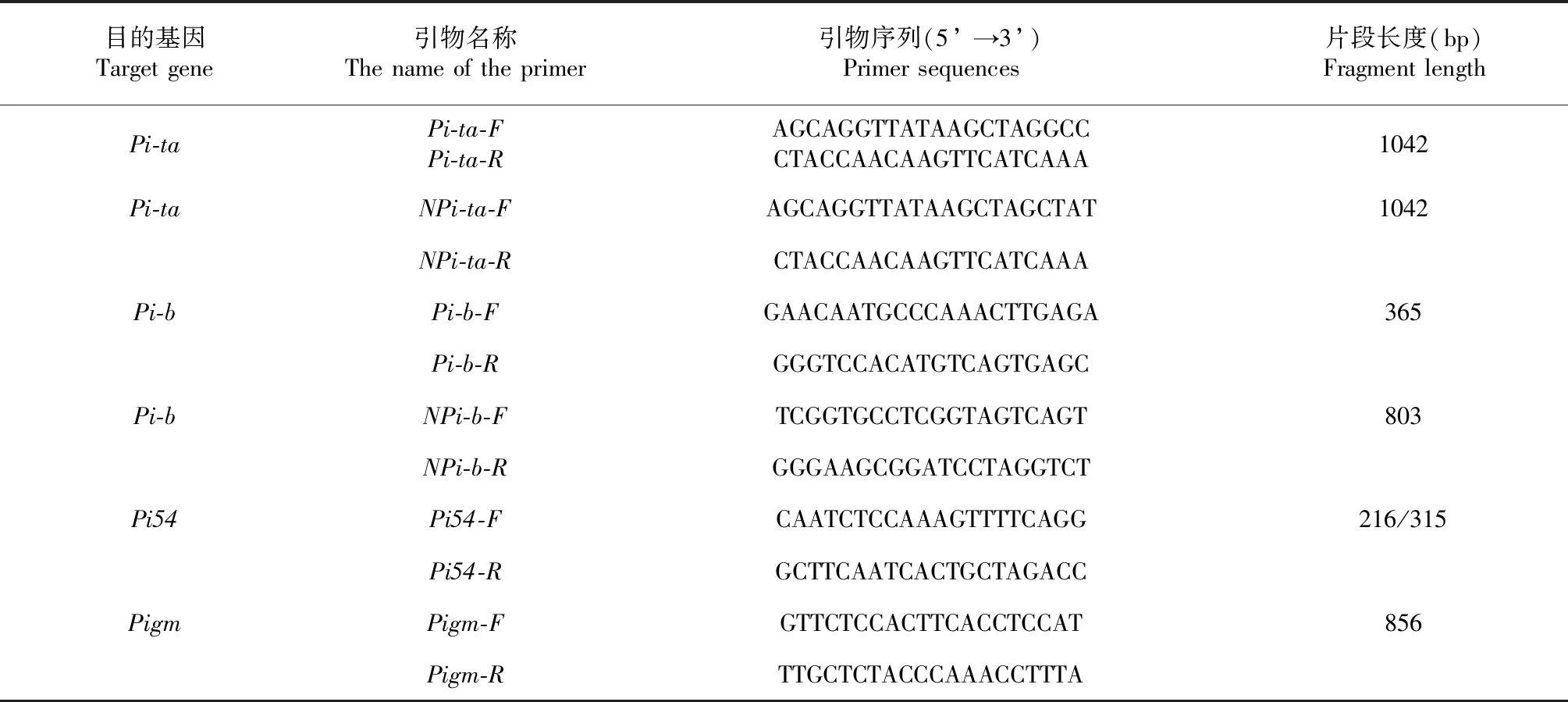
表1 PCR引物名称、序列及片段大小Table 1 Name, sequences and fragment size of specific primers used for PCR

表2 水稻品种穗颈瘟调查抗性分级和抗性评价标准Table 2 Investigation and evaluation criteria of resistance of rice varieties to neck blast
2.2 水稻穗颈瘟抗性鉴定
采用人工接种的方法,进行水稻穗颈瘟抗性鉴定。在2016年穗颈瘟接种鉴定中,80份试验材料中未发现0级免疫的材料,1级高抗的有38份,2级中抗的有21份,3级感病的有18份,4级高感的有3份。在2017年穗颈瘟接种鉴定中,80份试验材料中未发现0级免疫的材料,1级高抗的有22份,2级中抗的有22份,3级感病的有20份,4级高感的有16份。从连续两年的接种结果可以看出,抗病材料数量在逐渐减少,感病材料数量在逐渐增加。

图1 标记Pi-ta、Npi-ta、Pi-b、Npi-b、Pigm和 Pi54在部分品种中的PCR扩增结果Fig.1 The result of PCR using functional marker of Pi-ta, Npi-ta, Pi-b, Npi-b, Pigm, and Pi54 in part varieties
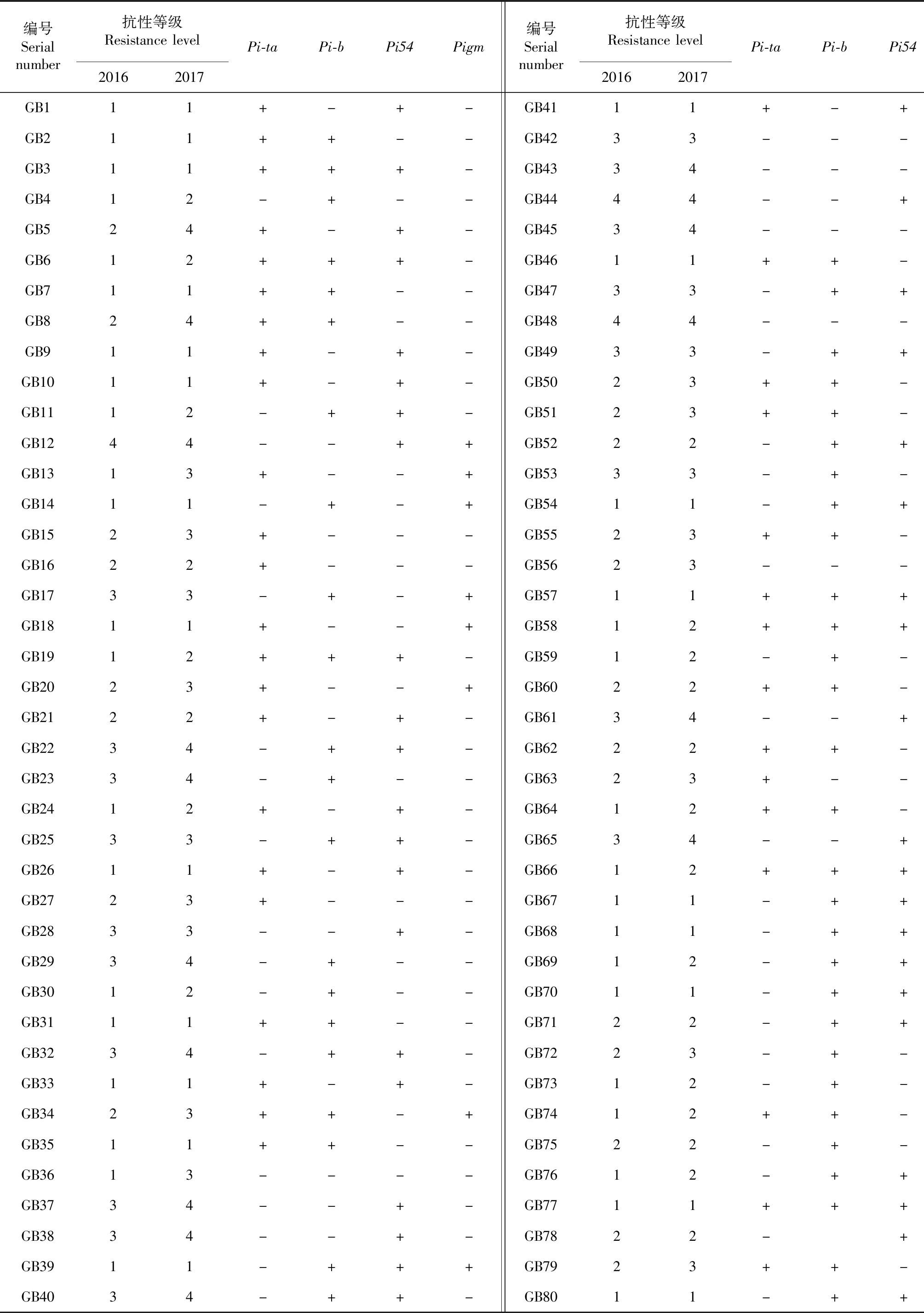
表3 80份试验材料穗颈瘟抗性鉴定与标记检测Table 3 Identification of resistance for panicle blast and marker detection for 80 test materials

表4 不同抗性基因组合的抗病性Table 4 Rice blast resistances of different genes and gene combinations
2.3 抗性基因分布与抗性相关性分析
通过对供试材料中4个抗性基因分布与抗性相关性分析表明(表4),80份试验材料中,两年穗颈瘟鉴定都表现为感病的有21份,检测到的抗性基因不超过2个,其中含Pi-b基因的材料有10份,含Pi54基因的材料有13份,没有检测到4个抗性基因的材料有4份。第1年接种鉴定表现为抗病,但第2年表现为感病的材料,总共有15份,除有3份材料没有检测到Pi-ta基因外,其余都检测到Pi-ta基因,表明2017年Pi-ta基因对稻瘟病抗性水平在下降。含3个抗性基因的材料有21份,其中两年表现均为抗病的有17份,其余4份第1年表现为抗病,第2年表现为感病,基因型为Pi-ta+Pi-b+Pigm。含有4个抗性基因的材料有1份,两年接种结果都表现为抗病。
3 讨 论
与传统的水稻抗病育种相比较,利用分子标记辅助选择育种,具有快速、简便、不受水稻生育期和环境影响的特点。范方军等[19]利用水稻抗病基因Pi-ta、Pi-b、Pi-km和Pi54的功能标记检测64份水稻品系,结果表明,抗稻瘟病基因Pi-ta与穗颈瘟存在显著相关性。戴小军等[22]利用水稻抗病基因Pi-ta、Pi-b、Pi9和Pi-km的分子标记,对抗性基因与抗性反应相关性进行探讨,结果表明,以Pi9为主效基因,同时聚合Pi-ta和Pi-b可以持久提高抗稻瘟病能力。王军等[23]利用4个抗稻瘟病基因分子标记,检测2007-2013年江苏省审定的粳稻品种,结果显示稻瘟病抗性基因Pi-ta、Pi-b、Pi-km与穗颈瘟的抗性呈正相关。向小娇等[24]研究发现水稻导入基因pi21,存在对产量的负效应,单独用来改良水稻品种的稻瘟病抗性,需要与抗性强的主基因聚合,通过多次回交和自交打破该基因与产量的不利连锁累赘。因此,在对基因选择聚合育种时,了解抗性材料的基因型背景,从而有针对性的进行多基因聚合[25-26]。并不是聚合基因越多越好,而是应该选择抗性好,抗谱互补的基因进行聚合。多基因的聚合不仅抗谱拓宽,而且对一些生理小种的抗性也提高,因为多基因的聚合不是简单地单个抗性基因的抗谱累加,而是抗性基因之间表现为极显著的基因互作[27]。
本研究中主要利用几个强致病稻瘟病小种,对80份资源材料进行连续两年穗颈瘟抗性鉴定,且利用水稻稻瘟病抗病基因Pi-ta、Pi-b、Pigm和Pi54的功能标记进行基因型分析,并分析不同基因型对穗颈瘟的抗性作用。在80份试验材料中,2016年接种鉴定有21份材料表现为感病,其都没有检测到Pi-ta基因,2017年接种鉴定有36份材料表现为感病,其中有12份检测到Pi-ta基因,说明2016年抗性基因Pita对江苏省内稻瘟病菌优势小种具有抗病性,2017抗性基因Pi-ta抗病能力在减弱。这是由于稻瘟病生理小种多、变化速度快、致病力差异大等原因,造成抗病材料在种植几年后,抗性就会逐渐丧失。两年接种鉴定都表现为高抗的材料有22份,其中基因型为Pi-ta+Pi54有5份,Pi-ta+Pi-b有5份,Pi-b+Pigm有1份,Pi-ta+Pigm有1份,Pi-ta+Pi-b+Pi54有3份,Pi-b+Pi54+Pigm有6份,Pi-ta+Pi54+Pigm有1份。结果表明,接种鉴定表现为高抗的材料,都是聚合2个及以上的抗病基因,且都含有主效抗病基因Pi-ta或Pigm。两年接种鉴定都表现为感病的材料有21份,其中基因型为Pi54+Pigm有2份,Pi-b+Pigm有2份,Pi-b+Pi54有6份,Pi-b有2份,Pi54有5份,没有检测到4种抗病基因的有4份。含有单个抗病基因的Pi-b和Pi54在抗性鉴定中基本已经丧失抗性。因此,应该有针对性的选择多基因进行聚合,仅仅聚合Pi54+Pigm、Pi-b+Pigm、Pi-b+Pi54仍表现为感病特征。虽然,单个抗病基因Pi-b和Pi54的抗性较弱,但基因型为Pi-ta+Pi-b+Pi54和Pi-b+Pi54+Pigm,两年接种抗性中都表现为抗病,一方面,稻瘟病抗性是由多个基因控制,其中主效基因对稻瘟病的抗性影响最大;另一方面,稻瘟病菌生理小种很多,不同抗性基因对生理小种的抗性存在差异。因此,在多基因聚合中,选择以Pi-ta或Pigm为主效基因,同时聚合Pi-b和Pi54基因,抗谱能够互补,可以提高抗稻瘟病的持久性。
4 结 论
从上述研究结果来看,针对复杂多变的稻瘟病菌种群结构,利用单一的抗性基因进行水稻抗病育种很难具有持久抗性,因此,在以后的水稻抗病育种中,需要引进不同的抗性资源,同时挖掘新的抗稻瘟病基因并进行多个有效抗病基因的聚合,才可能选育出具有广谱持久抗性的水稻新品种。
