桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
2020-04-20刘润红涂洪润梁士楚李月娟黄冬柳农娟丽
刘润红,陈 乐,涂洪润,梁士楚,姜 勇,*,李月娟,黄冬柳,农娟丽
1 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室,桂林 541006 2 兰州大学生命科学学院/草地农业生态系统国家重点实验室,兰州 730000 3 广西师范大学生命科学学院,桂林 541006
植物群落是由群落内共存物种在一定生境条件下相互依存、相互竞争和协同进化并与生境相互作用而形成的有机结合体[1- 2]。植物群落内物种间的相互关系作为种群生态学研究的核心问题,主要包括不同物种对群落中有限资源的竞争关系和不同种群间的稳定共存关系,而物种的生态位及种间联结特征可以很好地反映这些关系[3- 6]。其中,生态位是指群落中各种群所能利用的各种资源的总和及其与相关种群之间的功能关系,它体现了种群在群落中的地位、作用和重要性[2, 7]。而种间联结是指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[8- 9]。因此,研究群落内主要物种的生态位和种间联结特征不仅能够了解物种对资源的利用能力、物种间协作和竞争关系,而且有助于正确认识群落的结构、功能及演替规律,从而对植物资源的保护和利用、植被的恢复与重建以及生物多样性保护等具有重要的理论意义和应用价值[10- 11]。
桂林岩溶石山所处的中国西南喀斯特地区是世界三大集中连片喀斯特发育区中分布面积最大(55×104km2)、岩溶发育最典型、石漠化最严重、人地矛盾最尖锐,也是景观类型复杂、生物多样性丰富、生态系统极为脆弱的地区,在全球喀斯特生态系统中占据重要地位[12- 14]。长期以来,桂林岩溶地区由于地质背景特殊,景观异质性强,岩溶作用强烈,生态系统脆弱,抗干扰能力差,生态环境容量小,但人口众多,尖锐的人地矛盾导致植被破坏和水土流失日益严重,石漠化问题突出,严重制约了当地社会经济可持续发展,对该地区进行生态恢复与重建已刻不容缓[15- 16]。岩溶植被作为岩溶生态系统的主体,在维护岩溶生态系统稳定、调节区域碳平衡和保护物种多样性等生态服务功能方面具有重要的作用[17]。因此,植被的恢复与重建成为岩溶区石漠化治理和生态重建的首要任务。然而,在进行石漠化地区的植被恢复与重建过程中如何进行物种选择、配置和稳定植物群落的构建则是首先需要解决的问题。而解决这些问题的关键是弄清岩溶石漠化地区典型群落中植物的生物学和生态学特性。因此,研究岩溶石山典型植物群落中主要物种的生态位与种间联结,充分认识岩溶石山植被的组成、结构和演替过程中群落的动态特征,对岩溶石山退化生态系统的植被恢复和重建意义重大。
青冈(Cyclobalanopsisglauca)为壳斗科(Fagaceae)青冈属(Cyclobalanopsis)常绿乔木,具有较强的适应性,是岩溶生态系统顶极群落的建群种或共优种,对于维持岩溶生态系统的结构和功能具有重要的作用[18]。灌木层处于草本层与乔木层之间,是连接乔木和草本的纽带,故林下灌木层物种的替代作用和联结作用不容忽视[19]。青冈群落灌木层物种具有种类丰富、分布范围广泛、生命力强、萌生力强等特点,不仅在群落的演替过程中扮演着极其重要的角色,而且在区域生态环境保护方面也起着非常重要的作用。因此,研究桂林岩溶石山青冈群落灌木层物种的生态位与种间联结,对于岩溶地区植被恢复与重建具有重要的理论和实践意义。然而,以往对桂林岩溶石山青冈群落的研究主要集中在种群的结构特征[20]、种内种间竞争[21]以及群落数量分析[18]等方面,目前尚缺乏关于该群落灌木层生态位与种间联结方面的研究报道。鉴于此,本研究以桂林岩溶石山青冈群落灌木层主要物种为研究对象,在群落学调查的基础上,利用生态位宽度、生态位相似性、生态位重叠程度、方差比率法(VR)和基于2×2列联表的χ2检验、Pearson相关系数检验和Spearman秩相关系数检验等方法对桂林岩溶石山青冈群落灌木层主要物种的生态位与种间联结进行定量分析,以期回答以下3个科学问题:(1)桂林岩溶石山青冈群落灌木层主要物种的生态位特征如何?(2)桂林岩溶石山青冈群落灌木层主要物种的种间联结特征如何?(3)桂林岩溶石山青冈群落灌木层主要物种的生态位与种间联结有何关系?这些科学问题的合理解答,可以揭示该区域青冈群落灌木层主要物种的种间关系、现阶段群落组成的结构特点以及未来群落的动态、演替趋势和演替进程,进而为岩溶石山植被的恢复与重建、森林经营与管理、生物多样性保护等提供理论依据和实践指导。
1 材料与方法
1.1 研究区概况
研究区位于广西壮族自治区桂林市的典型岩溶地貌区,地理坐标为110°9′ —110°42′ E,24°40′ —25°40′ N,海拔多处于100—500 m之间。该地区地表形态复杂多样,岩溶地貌特征显著,地貌类型主要是由碳酸盐岩溶蚀为主形成的峰丛洼地和峰丛谷地,石峰密集,基岩裸露,地形复杂而破碎。该地属于中亚热带湿润季风气候,全年光照充足,气候温和,雨量充沛,年平均气温18—19℃,1月(最冷月)平均气温7.9℃,8月(最热月)平均气温28℃,全年无霜期309 d。年平均降雨量为1949.5 mm,降雨量年内分配不均,集中分布于4—7月,秋冬季干燥少雨;年日照时数为1670 h,年平均蒸发量为1490—1905 mm,年平均相对湿度为73%—79%,其中3—8月在80%以上,全年风向以偏北风为主,平均风速为2.2—2.7 m/s。研究区基岩主要为石灰岩,土壤以黄棕色或黑色石灰土为主,具有岩石裸露率高,土被不连续,土层浅薄,土壤钙含量高、含水量低等特点。受这些特殊的环境条件影响,该地区的适生植物具有耐旱性、石生性、嗜钙性等特点。在桂林岩溶石山地区,由于人为活动频繁和自然干扰,原生植被基本上被破坏殆尽,现存植被以次生的亚热带常绿落叶阔叶硬叶林为主。研究区内青冈群落层次结构较简单,乔木层以青冈为建群种,伴生种主要有齿叶黄皮(Clausenadunniana)、扁片海桐(Pittosporumplanilobum)、紫弹树(Celtisbiondii)、岩樟(Cinnamomumsaxatile)、菜豆树(Radermacherasinica)等;灌木层物种种类较丰富,其物种组成以青冈等乔木幼树,以及红背山麻杆(Alchorneatrewioide)、干花豆(Fordiacauliflora)、粗糠柴(Mallotusphilippinensis)、子凌蒲桃(Syzygiumchampionii)、山麻杆(Alchorneadavidii)、檵木(Loropetalumchinense)等灌木为主;草本层植物种类稀少,盖度较低,主要有三穗薹草(Carextristachya)、宽叶沿阶草(Ophiopogonplatyphyllus)、庐山香科科(Teucriumpernyi)、荩草(Arthraxonhispidus)等;此外,层间植物较丰富,主要有龙须藤(Bauhiniachampionii)、铁线莲(Clematisflorida)、香花崖豆藤(Calleryacinerea)等。
1.2 群落学调查
在充分实地踏查的基础上,于2017年7—9月在桂林岩溶石山受人为干扰较轻且发育完好的青冈群落分布区设置了4块样地,共建立了20个20 m×20 m样方,其中,阳朔县碑头村、桂林市秀峰区张家村、桂林市郊演坡山、阳朔县白沙镇的样方个数分别为7、3、5和5个。在此基础上,在每个样方的中心和四角,各设置1个5 m ×5 m的灌木样方,共100个。调查并记录灌木层内所有灌木个体(包括胸径≤2.5 cm的乔木幼树)的种名、株数、高度、基径和盖度等指标。同时,记录每个样方的经纬度、海拔、岩石裸露率、坡度、坡向和干扰程度等生境特征。
1.3 数据处理与分析
1.3.1重要值计算
为客观反映各物种在群落中的地位和作用,用重要值代表灌木层各物种的优势度,其计算公式如下[22]:
灌木层物种重要值(IV)=(相对多度+相对频度)/2
式中,相对多度=(某一物种在样方中的多度/所有物种多度总和)×100%,相对频度=(某一物种在样方中出现的频度/所有物种频度总和)×100%。
1.3.2生态位特征
以各个样方作为不同的资源位,用物种的重要值代表物种的资源利用状态,选取桂林岩溶石山青冈群落灌木层中重要值较高的20个物种(表1)进行生态位与种间联结分析。利用Levins指数(BL)[23]和Shannon指数(BS)[24]计算主要物种的生态位宽度,分别采用Schoener生态位相似性指数(Cik)[25]和Pianka生态位重叠指数(Oik)[26]测定主要物种间的生态位相似性和生态位重叠程度。
Levins生态位宽度(BL):
Shannon生态位宽度(BS):
式中,Pij=nij/Ni,nij为物种i在资源位j上的重要值,Ni为物种i在所有资源位上的重要值总和。Pij代表物种i在资源位j上的重要值占该物种在所有资源位上重要值总和的比例,r为样方总数。
Schoener生态位相似性指数(Cik):
式中,Cik为物种i和k的生态位相似性系数,Cik=Cki,值域为[0,1],其值越大表示生态位相似程度越高;Pij和Pkj分别是物种i和k在资源位j上的重要值。
Pianka生态位重叠指数(Oik):
式中,Oik为物种i和k的生态位重叠指数,值域为[0,1],其值越大表示生态位重叠程度越高;Pij和Pkj分别是物种i和k在资源位j上的重要值。
1.3.3总体联结性检验
采用Schluter[27]提出的方差比率法(VR)来测定灌木层主要物种间的总体联结性,并利用统计量W来检验总体联结是否显著。其计算公式如下:
Pi=ni/N
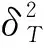
1.3.4种间联结性分析
采用χ2统计量对种间联结性进行定性研究,以确定实测值与在机率基础上预期值之间偏差的显著程度。鉴于本研究为非连续性取样,因此χ2统计量需采用Yates的连续校正公式来纠正[28]。公式如下:
式中,N为总样方数,a为2个物种同时出现的的样方数,b、c分别为2个物种单独出现的样方数,d为2个物种都不出现的样方数。种间联结性通常分为正联结和负联结2类,设V=((a+d)-(b+c))/(a+b+c+d),当ad>bc时,则V>0,种间联结性为正联结;当ad
χ2检验仅定性判断了种间联结性是否显著,并不意味着那些χ2检验不显著的种对间就不存在联结性,且无法区分联结强度的大小,不能清楚地表达物种间联结性的差异性[29]。而Pearson相关检验和Spearman秩相关检验则是基于数量数据,能比较准确客观地反映物种间的线性关系,从而能对χ2检验进行有效的补充和完善。因此,在进行种间联结χ2检验后,需进一步采用Pearson相关检验和Spearman秩相关检验确定其联结程度。鉴于此,本研究利用灌木层主要物种的重要值作为Pearson相关检验和Spearman秩相关检验的数量指标,对种对间的线性关系进行定量分析,分析它们同时出现的可能性。
Pearson相关系数计算公式如下:
Spearman秩相关系数计算公式如下:

此外,为了解生态位与种间联结的相关性,对桂林岩溶石山青冈群落灌木层主要物种间的Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠值进行回归分析。分别采用Excel 2019和R 3.5.2计算重要值和生态位与种间联结特征。采用种间联结分析程序包spaa[30]中的niche.width()、niche.overlap()、sp.assoc()、sp.pair()函数分别计算生态位宽度、生态位相似性和生态位重叠程度、χ2检验、Pearson相关和Spearman秩相关检验结果,通过psych包中的corr.test()函数进行Pearson相关和Spearman秩相关显著性检验。
2 结果与分析
2.1 重要值特征
在所调查的100个灌木样方中,灌木层共有植物88种,隶属于37科73属。如表1所示,灌木层20个主要物种中,平均重要值排序为青冈(42.26)>红背山麻杆(8.80)>干花豆(6.56)>齿叶黄皮(4.84)>粗糠柴(3.76)>紫弹树(3.16)>扁片海桐(2.46)>子凌蒲桃(2.35)>山麻杆(2.17)>千里香(1.92)>檵木(1.91)>胡颓子(1.39)>网脉山龙眼(1.24)>樫木(1.11)>菜豆树(1.06)>合欢(1.00)>白萼素馨(0.94)>白皮乌口树(0.73)>樟叶槭(0.67)>石岩枫(0.65)。
2.2 生态位宽度
由表1可知,桂林岩溶石山青冈群落灌木层主要物种的生态位宽度存在较大差异,BL和BS的变化范围分别为2.72—72.55和1.50—4.37,其中青冈的生态位宽度最大,其BL和BS分别为72.55和4.37,樫木的生态位宽度最小,BL和BS为2.72和1.50,多数物种的BL和BS生态位宽度分别在10—40和2—4之间。Levins和Shannon生态位宽度测定结果虽排序稍有差异,但总体上基本一致。重要值较大(小)的物种,其生态位宽度一般较大(小),但两者排序并非完全一致,如红背山麻杆重要值较干花豆大而生态位宽度却较干花豆小,网脉山龙眼和樫木的重要值虽然处于中等水平,但是两者的生态位宽度却比较小。

表1 桂林岩溶石山青冈群落灌木层主要物种的重要值和生态位宽度值
2.3 生态位相似性
由表2可知,青冈群落灌木层物种的生态位相似性(Cik)介于0—0.495之间,Cik最大的是青冈和干花豆(0.495),在0.3以上的有20个种对(占10.53%),处于0.2—0.3的有38个种对(占20.00%),介于0—0.2的有120个种对(占63.16%),为0的有12个种对(占6.31%)。生态位宽度较大的青冈和干花豆的生态位相似性达0.495,而生态位宽度较窄的樫木和樟叶槭的生态位相似性为0,说明生态位相似性与生态位宽度有一定关联。从表2还可看出,青冈和干花豆生态位相似性高达0.495,表明青冈和干花豆幼树对环境资源的需求比较相似。而子凌蒲桃和胡颓子,子凌蒲桃和樫木,山麻杆和胡颓子,山麻杆和樫木,檵木和胡颓子,檵木和樫木,胡颓子和菜豆树,网脉山龙眼和石岩枫,樫木和菜豆树,樫木和樟叶槭,合欢和白萼素馨,合欢和石岩枫的Cik均为0,它们均没有在同一资源位中出现,说明它们对环境资源的需求基本不同。青冈群落灌木层主要物种的生态位相似性平均值为0.156,且多数种对的生态位相似性在其平均值附近,生态位相似性较小,表明大部分物种对环境资源的要求差异较大。
2.4 生态位重叠程度
由表3可知,青冈群落灌木层物种的Pianka生态位重叠指数(Oik)介于0—0.663之间,Oik>0的有178个种对,占总种对数的93.68%,其中Oik<0.5的种对有185对,占总种对数的97.37%,Oik>0.5的种对有5对,占总种对数的2.63%。未产生生态位重叠(Oik=0)的种对有12对,占总种对数的6.32%,各物种间普遍存在生态位重叠,表明这些物种对环境资源的利用较充分,种间关系协调。灌木层主要物种之间的生态位重叠程度普遍较低,生态位重叠平均值为0.173,表明灌木层主要物种之间竞争不激烈,种间关系较稳定。生态位宽度较小的物种间也存在生态位重叠程度较高的种对,如胡颓子和网脉山龙眼、扁片海桐和菜豆树的生态位宽度较小,但是它们的生态位重叠程度却较大,由此可见生态位宽度与生态位重叠程度之间并不存在绝对的正相关关系。生态位相似性较高(低)的种对,它们的生态位重叠程度一般较大(小),如青冈和干花豆,子凌蒲桃和檵木的生态位相似性分别为0.495和0.491,其生态位重叠值分别为0.503和0.663。另外,生态位宽度较大的物种与群落中其他物种产生生态位重叠的概率较大,如生态位宽度最大的青冈与其他物种发生生态位重叠的概率为100%,可能是因为该物种地理分布范围广,资源利用能力强,能与许多生态位较窄的物种产生生态位重叠。
2.5 总体联结性分析

2.6 种对间的联结性分析
χ2检验结果显示(图1和表5),在灌木层20个主要物种构成的190个种对中,呈正联结的种对有103对,占总种对数的54.21%,其中,呈极显著、显著和不显著正联结的种对分别有4、14和85对,分别占总种对数的2.10%、7.37%和44.74%;呈负联结种对有87对,占总种对数的45.79%,其中,呈极显著、显著和不显著负联结的种对分别有1、5和81对,分别占总种对数的0.53%、2.63%和42.63%;正负联结种对的比值为1.18,正联结占优势,这与灌木层总体联结性呈正联结的结果一致。显著联结种对有24对,检验显著率为12.63%;不显著联结种对有166对,占总种对数的87.37%,种对间联结较弱,各物种间呈独立分布格局。
Pearson相关检验结果显示(图2和表5),呈正相关的种对有82对,占总种对数的43.16%,其中,呈极显著、显著和不显著正相关的种对分别有14、7和61对,分别占总种对数的7.37%、3.68%和32.11%;呈负相关种对有108对,占总种对数的56.84%,其中,呈极显著、显著和不显著负相关的种对分别有7、8和93对,分别占总种对数的3.68%、4.21%和48.95%;正负相关种对的比值为0.76,显著相关种对有36对,检验显著率为18.95%;不显著相关种对有154对,占总种对数的81.05%,绝大部分种对为不显著相关,相关性不强,这同χ2检验的结果基本一致。



表4 桂林岩溶石山青冈群落灌木层主要物种间的总体联结性

图1 桂林岩溶石山青冈群落灌木层主要物种间χ2检验半矩阵图 Fig.1 Semi-matrix diagram of interspecific association χ2 test among main species in shrub layer of Cyclobalanopsis glauca community in karst hills of Guilin
Spearman秩相关检验结果显示(图2和表5),呈正相关的种对有96对,占总种对数的50.53%,其中,呈极显著、显著和不显著正相关的种对分别有25、10和61对,分别占总种对数的13.16%、5.26%和32.11%;呈负相关种对有94对,占总种对数的49.47%,其中,呈极显著、显著和不显著负相关的种对分别有12、7和75对,分别占总种对数的6.32%、3.68%和39.47%;正负相关种对的比值为1.02,显著相关种对有54对,检验显著率为28.42%;不显著相关种对有136对,占总种对数的71.58%,种对间相关较弱,各物种间呈独立分布格局,这同χ2检验的结果基本一致。
2.7 种间联结与生态位相似性和生态位重叠的回归分析
对桂林岩溶石山青冈群落灌木层主要物种间的Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠值进行回归分析,结果如图3所示。桂林岩溶石山青冈群落灌木层主要物种间的Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠值之间均呈极显著正相关(P<0.001),即种间正联结越强,生态位相似性和生态位重叠程度越大,反之,负联结越强,生态位相似性和生态位重叠程度越小。
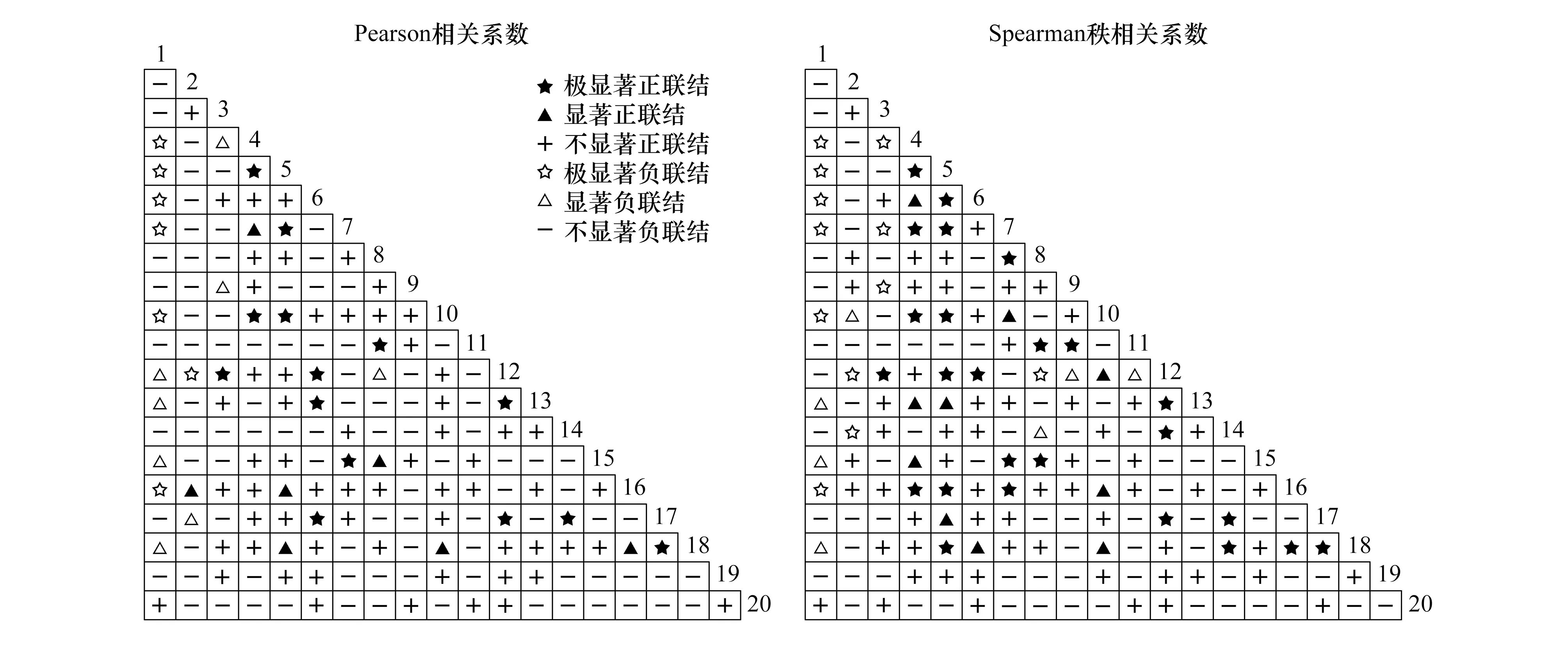
图2 桂林岩溶石山青冈群落灌木层主要物种间相关半矩阵图Fig.2 Semi-matrix diagram of interspecific correlation among main species in shrub layer of Cyclobalanopsis glauca community in karst hills of Guilin
表5 桂林岩溶石山青冈群落灌木层主要物种的χ2检验、Pearson相关检验和Spearman秩相关检验结果比较
Table 5 The comparison ofχ2test coefficient, Pearson′s correlation coefficient and Spearman′s rank correlation coefficient among main species in shrub layer ofCyclobalanopsisglaucacommunity in karst hills of Guilin

检验方法Test methods正联结(相关)Positive association(correlation)负联结(相关)Negative association(correlation)极显著Distinctlysignificant(P≤0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum极显著Distinctlysignificant(P≤0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sumχ2检验χ2 test4(2.11)14(7.37)85(44.74)103(54.21)1(0.53)5(2.63)81(42.63)87(45.79)Pearson相关Pearson's correlation14(7.37)7(3.68)61(32.11)82(43.16)7(3.68)8(4.21)93(48.95)108(56.84)Spearman秩相关Spearman's rank correlation25(13.16)10(5.26)61(32.11)96(50.53)12(6.32)7(3.68)75(39.47)94(49.47)

图3 桂林岩溶石山青冈群落灌木层主要物种间Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠的回归分析Fig.3 Regression analysis between Pearson correlation coefficient, Spearman rank correlation coefficient and niche similarity and niche overlap among main species in shrub layer of Pterocarya stenoptera community in karst hills of Guilin
3 讨论
3.1 生态位宽度特征
生态位宽度是一个种群所能利用的各种资源的总和,它很好地反映了植物种群的生态适应性和对环境资源的利用能力[31]。生态位宽度大小取决于物种的种间竞争力、分布范围和生态适应性等因素。一般而言,物种的生态位宽度越大,则它对环境的适应能力越强,对资源的利用更充分,分布范围越广泛[32]。通常认为物种的生态位越宽,其特化程度越小,更倾向于泛化种,常是群落中的优势物种[33]。本研究表明,青冈、红背山麻杆和干花豆等物种的生态位宽度较大,说明它们具有较强的资源利用和环境适应能力,分布范围较为广泛,为桂林岩溶石山青冈群落灌木层的优势种或伴生种,对维持群落内部环境以及生态环境的稳定起重要作用。而樟叶槭和石岩枫等物种的生态位宽度较窄,说明它们对环境的适应以及资源的利用能力相对较弱,对环境变化比较敏感,地理分布范围较窄,空间分布不均匀,适宜的生境较少,更倾向于特化种,在群落中的竞争力明显较弱,在群落的发展过程中存在被淘汰的危险。因此,在对桂林岩溶石山青冈群落进行经营管理时,应避免对这些种群所在的生境进行干扰。
重要值和生态位宽度都是度量物种在群落中的地位和作用的综合指标,但这两者的生态学意义不尽相同,重要值体现了物种在群落中的优势程度,代表了物种在群落中的地位和重要性,而生态位宽度则反映了植物种群对环境资源的利用能力及其生态适应性等生态位特征[33]。许多研究表明,物种的重要值越大(小)则其生态位宽度一般也越大(小)[34]。本研究发现群落内大部分物种的重要值越大(小),其生态位宽度一般也越大(小),但也有例外,如红背山麻杆的重要值较干花豆大而生态位宽度却较干花豆小,网脉山龙眼和樫木的重要值虽然处于中等水平,但是两者的生态位宽度却较小,这说明重要值并不是影响生态位宽度的唯一因素,生态位宽度的大小与其分布频度也密切相关,分布频度越大,生态位宽度也越大[35]。而网脉山龙眼和樫木等物种,资源利用和环境适应能力较弱,在资源位中出现的次数少,分布范围较窄,故它们的生态位宽度较小。
3.2 生态位相似性特征
生态位相似性是衡量物种间资源利用相似程度的重要指标,可以利用其判断物种间是否存在竞争以及群落是否稳定[36]。青冈和干花豆等种对的生态位相似性程度较高,表明它们的生活习性相近,对环境资源的需求比较相似,故它们的生态位相似性程度也较高。而子凌蒲桃和胡颓子等12个种对的Cik值均为0,这可能是由于这些物种对环境资源的利用和要求基本不同。许多研究表明,生态位相似性与其生态位宽度呈正相关关系,生态位宽度较大(小)的物种间,其生态位相似性程度一般也较高(低)[7]。本研究也发现,生态位宽度较大的青冈和干花豆的生态位相似性达0.495,而生态位宽度较窄的樫木和樟叶槭的生态位相似性却为0。但是生态位宽度较小的物种间,由于它们的生物学、生态学特性相似,可能会产生较大的生态位相似性[37]。如,本研究中的樫木和白萼素馨的生态位宽度均较小,但是它们之间的生态位相似性却较大。这与刘润红等[33]对漓江河岸带枫杨群落主要木本植物种群的生态位的研究结果一致,但是青冈群落灌木层主要物种的生态位相似性平均值为0.156,远低于漓江河岸带枫杨群落中灌木层主要物种的生态位相似性,表明青冈群落灌木层主要物种对资源利用差异较大,物种间潜在的竞争较少,种间关系相对稳定。此外,桂林岩溶石山生境复杂多样,生境过滤作用导致物种多集中分布于适合的生境斑块中,故某些物种虽然生态位宽度较小,但是它们的生态位相似性却较高。
3.3 生态位重叠特征
生态位重叠是指两个或两个以上生态位相似的物种生活于同一空间时分享或竞争共同资源的现象[38]。生态位重叠是反映物种对环境资源利用能力差异性和竞争关系的重要指标,生态位重叠程度越大表明物种之间的生活型越相似,对环境资源的生态需求越相似,物种之间可能存在激烈竞争[36]。本研究发现桂林岩溶石山青冈群落灌木层主要物种之间的生态位重叠程度普遍较低,生态位重叠平均值为0.173,表明灌木层主要物种之间对环境资源的利用和要求差异较大,物种间竞争不激烈,种间关系较稳定。通常,生态位宽度较大的物种,其资源利用能力较强,分布范围较广,因而与其他物种间的生态位重叠程度也较大,而生态位较窄的物种间,其生态位重叠程度比较小[25]。但是由于生态位宽度较大的物种本身的生物生态学特征不一定完全相同,对环境资源的利用和要求并非完全一致,使得生态位宽度大(小)的物种间也可能会出现较低(高)幅度的生态位重叠,如胡颓子和网脉山龙眼等物种的生态位宽度较小,但是它们的生态位重叠程度却较大。这可能与桂林岩溶石山恶劣的生境条件有关,该地区岩石裸露率高,土层浅薄,地形破碎,土被不连续,土壤含水量和土壤养分等资源匮乏,导致物种间产生资源利用竞争,某些植物为共享稀缺的环境资源,生态位宽度相对压缩,多倾向于聚集在局部适宜的生境斑块中,而在适宜生境斑块以外的空间分布较少,以致虽然这些物种的生态位宽度较小,但是却产生较大程度的生态位重叠。这与张忠华等[39]和钟军弟等[40]认为这可能是由喀斯特生境的高异质性所引起的研究结论相吻合。
3.4 种间联结特征
种间联结是各个物种在不同生境中相互影响、相互作用而形成的有机关系,体现了由于群落生境差异而造成的不同物种间的相互联系[6]。种间联结性反映了物种间的相互影响和群落动态,物种间的总体联结体现了群落演替的进程及其稳定性,而群落稳定性是种间联结性的表达方式[41]。一般来说,处于演替早期的群落,物种间的联结程度较低,正负联结比值偏小,群落处于不稳定阶段。随着群落的演替更新,种间联系也逐渐接近于正联结,群落正负联结比值变大,群落稳定性也逐渐增强,群落向顶极方向演替[42]。桂林岩溶石山青冈群落灌木层主要物种的种间联结测定结果表明,灌木层主要物种间的总体联结性表现为显著正联结,这与韩文衡等[43]对桂西北喀斯特地区常绿落叶阔叶混交林优势物种的种间联结性研究结果一致,说明桂林岩溶石山青冈群落灌木层处于相对稳定阶段,群落结构及其种类组成将逐渐完善和稳定,接近群落演替发展的后期阶段。综合χ2检验、Pearson相关检验和Spearman秩相关检验结果发现,青冈群落灌木层主要物种多呈不显著联结,种对间联结程度较弱,独立性相对较强,这与涂洪润等[44]对桂林岩溶石山青冈群落乔木层优势种的种间关联性分析结论一致。这可能是由于桂林岩溶石山物种的种间关系主要受到环境限制和种间相互作用这两个方面的影响。其中,环境限制主要体现在岩溶石山生境异质性上,岩溶石山地形破碎、石漠化严重、岩石裸露率高、成土条件差、土层浅薄、土壤贫瘠,且该地区多暴雨,雨水冲刷能力强,造成水土流失严重,地表水源涵养和调蓄能力弱等恶劣地形地貌特征,导致即使在同一地段、坡位或坡向的小生境,其生境差异也十分明显,形成了不连续、不均匀和高度复杂的大量异质生境斑块[16]。在种间关系方面,岩溶石山地区大量复杂多样的异质生境斑块强化了种内和种间竞争关系,不同生态适应性的植物利用不同的空间和资源,物种占据着不同的生态位,种间关系发生较大改变,物种处于相对独立状态,造成负联结种对数较多[45]。因此,岩溶石山地区的生境异质性对植物种间关系的形成具有重要的影响,导致了物种对生境的选择,并在不同生境斑块下生态位发生改变,呈负联结的种对数增多,使得青冈群落灌木层中物种间的正负联结比基本都小于1,种对间联结程度不明显,独立性相对较强。
3.5 生态位与种间联结的相关性
物种的种间联结性与其生态位相似性和生态位重叠程度联系密切:种对间的正联结反映了植物利用资源的相似性和生态位的重叠性;负联结表明了物种间的相互排斥性和生态位分离[46]。本研究发现桂林岩溶石山青冈群落灌木层主要物种间的Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠值之间均呈极显著正相关,总体上表现为种间正联结越强,生态位相似性和生态位重叠程度越大,负联结越强,生态位相似性和生态位重叠程度越小。这一结论在许多关于生态位和种间联结的研究中也得到证实[5, 46- 47]。正联结的种对间,其生态位重叠值程度较大,反映出这些物种对生境要求的一致性[48],如粗糠柴和齿叶黄皮,为显著正联结种对,其生态位相似性和生态位重叠较大,分别为0.480和0.507,这是因为这2个物种均为喜阳物种,有着相似的资源利用策略。但是青冈与红背山麻杆、干花豆等物种呈显著或不显著负联结,其生态位重叠值却较高。由此可见,种间负联结越强,其生态位重叠值不一定会越小,这可能是因为种对间的联结性不仅与物种生态位存在一定的相关性,也与物种对分布的频率、生境和资源利用能力等条件有关[49]。此外,种对间的负联结成因比较复杂,由生境差异导致的负联结,种对间的生态位重叠程度较低,而由资源竞争导致的负联结,种对间的生态位重叠程度较高[50]。桂林岩溶石山生境恶劣,青冈与红背山麻杆、干花豆等物种对资源竞争激烈,导致它们呈显著或不显著负联结,生态位重叠程度较高。
3.6 植被保护与恢复策略
从桂林岩溶石山青冈群落灌木层主要物种的生态位与种间联结特征来看,在对桂林岩溶石山进行植被恢复与重建实践过程中,在物种选择上,应参照该地区当前恢复阶段的天然群落,在了解物种生物学、生态学特性的基础上,有选择性地将生物学特点和生态学习性相似,对生境具有相似要求的物种搭配在一起,选择环境适应能力强,即生态位宽度较大的物种作为先锋种(如青冈和山麻杆等),选择生态位重叠程度较小的物种作为伴生种,多选择种间正联结较强的物种进行搭配种植,防止种间产生恶性竞争,将不同的种群配置在群落的相应空间位置,充分合理利用有限的环境资源,使群落处于一种互补互利的协调关系,从而维护群落的稳定性和多样性,促进群落可持续发展。此外,本研究区内青冈群落多处于演替的中晚期,群落中的青冈主要以大树为主,阴蔽的林下青冈幼苗较多,天然更新能力较好,但在其向中高龄阶段发展的过程中,因群落内部的竞争作用等因素影响,青冈幼龄个体死亡率较高,导致其种群更新缓慢。由于青冈为阳生树种,因此,在自然封育过程中,应对郁闭度较高的群落采取适度的人工抚育措施,如适当地间伐、砍灌、清理群落内的衰老木、枯死木以及风倒木,建立小面积林窗,增大林内的辐射强度,同时加强对中小径级青冈的保护,为其提供良好的生长环境,以提高青冈幼苗存活率,促进青冈幼苗和幼树的生长和发育,从而诱导青冈群落的演替更新,推动岩溶石山植被的恢复与重建,改善该地区生态系统结构与功能。
