马铃薯光周期敏感性研究
2020-04-10马永珍王芳王舰
马永珍 王芳 王舰


摘要:为了研究不同光周期处理对马铃薯生长的影响及光敏性,以国外2号、闽薯1号和青薯9号3个马铃薯品种为试验材料,设定3种不同光周期分别为8、12、16 h/d,通过分析马铃薯的表型性状(株高、茎粗、茎节数、叶片数、根长、根数、鲜质量、干质量)和生理生化指标(可溶性蛋白含量、过氧化氢酶活性、超氧化物歧化酶活性、脯氨酸含量、丙二醛含量)筛选出能够反映马铃薯光周期敏感性的指标性状,对3个马铃薯品种的光周期相对敏感性进行分析。结果表明:3个马铃薯品种表型性状鲜质量、干质量、茎粗和根长均随光照时数的延长而增加。方差分析表明,光周期对马铃薯的干质量、根数有极显著影响(P<0.01),对鲜质量以及总根长有显著影响(P<0.05);品种对株高和根数有极显著影响(P<0.01);光周期与品种互作效应对茎粗、可溶性蛋白含量、过氧化氢酶(CAT)活性、超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量和脯氨酸(Pro)含量均有极显著影响(P<0.01)。筛选干质量、鲜质量、根数和根长4个表型性状作为光周期相对敏感度指标,计算得出青薯9号相比闽薯1号和国外2号为光周期不敏感材料。
关键词:马铃薯;光周期;表型性状;生理生化指标;敏感性分析
中图分类号: S532.01;S532.032 文献标志码: A 文章编号:1002-1302(2020)03-0106-06
马铃薯(Solanlum tuberosum L.)是茄科(Solanaceae)茄属(Solanum)双子叶多年生草本植物,是继水稻、小麦、玉米第四大世界粮食作物,也可作为蔬菜,同时马铃薯还是化工原料和动物饲料的重要原料[1-2]。马铃薯喜光、喜冷凉性环境,对光周期和温度反应的敏感性是影响其资源引进能否成功的重要环境因子[3],研究和科学评价马铃薯对光周期反应的敏感性,为开展马铃薯光周期敏感性及其相关性状的遗传研究奠定基础。
目前,相关光周期的研究表明,延长光照时间可以增加浮萍干物质的积累[4],增加茄子幼苗的株高、茎粗和生长速率还能通过提高过氧化物酶(POD)和过氧化氢酶(CAT)活性来增强抗逆性[5]。缩短光照时间会使苦瓜POD活性和可溶性蛋白质含量升高,CAT和超氧化物歧化酶(SOD)活性降低[6]。番茄幼苗生长发育难以适应短日照光周期,对番茄生长发育处理效果最好的是1 ∶ 1红蓝光配比的 14 h/10 h处理,光同期是影响番茄小苗生长发育的主要因素[7]。菜豆短日照(SD)处理能够促进开花,抑制营养生长;长日照(LD)则相反[8]。模拟8、12、16 h/d 3种光周期处理金线莲后发现,短日照8 h/d处理使初花期延迟8 d,而长日照16 h/d处理使初花期提前3 d,且成花率较高[9]。不同的光周期条件下,光周期反应的敏感时期不同,如玉米自交系CML28,短日照9 h/d处理下7叶期为光周期反应敏感的时期,长日照15 h/d处理下9叶期为光周期反应敏感的时期[10]。谷子农艺性状光周期相对敏感程度依次为穗数>抽穗期>穗长>株高>叶片数>穗粒质量,以穗数、穗长和抽穗期为光周期敏感指标[11]。对马铃薯的研究表明,短光周期培养有利于试管薯的诱导,但产生的试管薯较小,适当地延长光照时间有利于诱导较大的试管薯[12]。青薯9号壮苗培养30 d后,在无激素添加的MS固体培养基上10 h/d光照周期试管薯结薯效果最好,平均结薯1个/株,单瓶结薯率为92%[13]。在光照12 h、黑暗12 h处理下,弗乌瑞它微型薯植株徒长,净光合速率、气孔导度等指标升高,且块茎干物质及淀粉含量最高,而还原糖含量最低;光照16 h、黑暗8 h处理下,产量最高,薯块大,平均单薯质量最大[14]。
本研究模拟3种不同的光周期,观察和分析光周期对马铃薯表型性状和生理生化指标的影響,并利用方差分析3种不同光周期处理下8个表型性状和5个生理生化指标的差异,筛选出光周期相对敏感度指标,评价3个马铃薯品种光周期敏感性,为开展马铃薯光周期敏感性及其相关性状的遗传研究奠定基础。
1 材料与方法
1.1 试验材料、仪器与试剂
1.1.1 试验材料 试验材料为培养30 d的青薯9号(Q9)、闽薯1号(M1)和国外2号(G2号)组培苗,由青海省农林科学院生物技术研究所提供。
1.1.2 试验仪器 智能光照培养箱(MGC-300A),购自上海一恒科技有限公司;冷冻离心机(Bio-rad),购自上海凌仪生物科技有限公司;恒温水浴锅(THZ-82),购自常州国华电器有限公司;紫外可见风光光度计(T6新世纪),购自北京普析通用仪器有限责任公司;电子精密天平(PL203),购自梅特勒-托利多仪器(上海)有限公司;漩涡混合器(XW-80A),购自上海精科实业有限公司;-80 ℃超低温冰箱(Forma 900),购自赛默飞世尔(苏州)仪器有限公司;高通量动植物组织研磨机(SPEX Geno/Grinder 2010),购自美国SPEX SamplePrep公司;丰量牌电子数显游标卡尺(GB/T 1214.2—1996《游标类卡尺 游标卡尺》)、直尺(20 cm)等。
1.1.3 试剂 植物可溶性蛋白(Sp)定量测定试剂盒(A045-2)、过氧化氢酶(CAT)测试盒(A007-1)、超氧化物歧化酶(SOD)测试盒(A001-1)、丙二醛(MDA)测试盒(A003-1)和脯氨酸(Pro)测试盒(A107-1-1)均购自江苏省南京建成生物工程研究所,浓硫酸由笔者所在实验室提供。
1.2 试验处理
试验于2018—2019年在青海大学生物技术研究所实验室内进行。试验材料除顶,剪取2~3 cm带叶茎段,接种MS培养基(pH值6.0),置于光照时长为8、12、16 h/d条件下,光照度为4 950 lx,温度昼夜分别为18、16 ℃,湿度保持75%左右,每瓶接种5株,每个处理接种42瓶,处理35 d后取样,3次重复,叶片混装在1.5 mL离心管中用于测定不同处理下的生理生化指标。
1.3 试验方法
(1)表型性状测定。株高、茎粗、茎节数、叶片数、总根长和根数直接观察测量。(2)植株干鲜质量测定。将试管苗用无菌水清洗,吸去表面的水分,称量鲜质量;然后将植株放入锡箔纸中于105 ℃杀青40 min,85 ℃烘干至恒质量,称干质量[15]。(3)生理生化指标测定。Sp含量测定采用考马斯亮蓝法[16];CAT活性测定采用可见光法[16];SOD活性测定采用羟胺法[16];Pro含量测定采用酸性茚三酮法[16];MDA含量测定采用硫代巴比妥酸(TBA)法[16]。所有表型性状和生理生化指标重复测定3次,结果取平均值。
1.4 数据分析
数据处理与方差分析均采用均Excel 2007和DPS 9.50软件进行处理。
2 结果与分析
2.1 不同光周期处理对不同马铃薯品种组培苗表型性状的影响
由图1、表1可知,不同光周期处理下不同马铃薯品种组培苗之间的表型性状表现是不同的。G2、M1和Q9的鲜质量和干质量均随着光照时间的延长而增加,表明延长光照时间有利于马铃薯组培苗的生长,同时3个马铃薯品种的鲜干质量均表现为Q9>M1>G2。G2的株高在8 h/d处理下显著高于12、16 h/d处理(P<0.05);M1的株高在12 h/d处理下显著低于8、16 h/d处理(P<0.05),且在 16 h/d 处理下最大;Q9的株高在不同光周期处理下差异不显著,但在16 h/d处理下达到最大,表明短日照处理有利于G2伸长生长,而长日照处理有利于M1和Q9的伸长生长。G2、M1、Q9的茎粗也随着光照时间的延长而增加,表明长日照处理有利于马铃薯组培苗茎粗的生长,也就是说长日照处理有利于培育马铃薯壮苗。G2的叶片数在不同光周期处理下没有显著差异,但在16 h/d处理下达到最大;M1的叶片数在16 h/d处理下显著高于8、12 h/d 处理(P<0.05);Q9的叶片数在12 h/d处理下显著高于8、16 h/d处理(P<0.05),表明长日照处理有利于G2和M1叶的生长,而在日中性下有利于Q9叶的生长。G2、M1和Q9茎节数均在 16 h/d 处理下达到最大,且M1在12 h/d处理下显著低于8、16 h/d处理(P<0.05)。G2、M1和Q9的根长均随着光照时间的增加而延长。G2和Q9的根数均在12 h/d处理下达到最大,M1的根数随着光照时间的延长而增加。表明日中性处理有利于G2和Q9根的生长,长日照处理有利于M1根的生长。
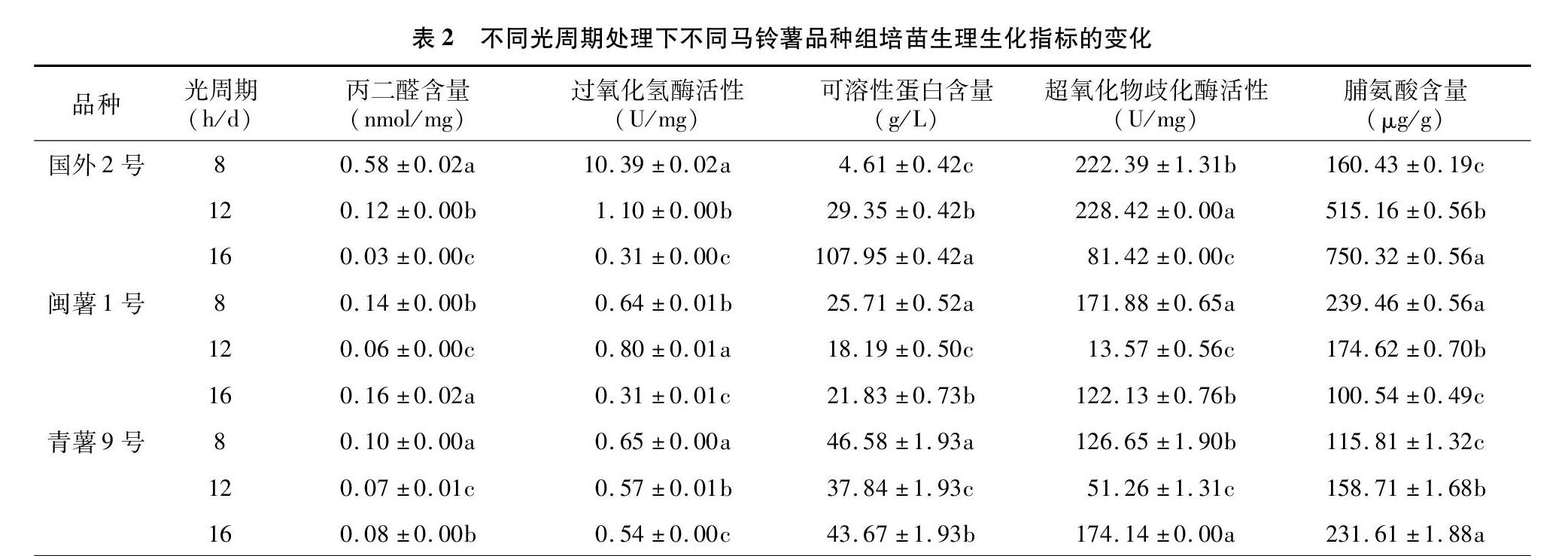
2.2 不同光周期处理对不同马铃薯品种生理生化指标的影响
由表2可知,不同光周期对不同马铃薯品种生理生化指标的影响是不同的。马铃薯G2和Q9的MDA含量均在8 h/d处理下显著高于12、16 h/d处理(P<0.05),M1在16 h/d处理下显著高于8、12 h/d 处理(P<0.05),表明短日照处理有利于G2和Q9 MDA含量的提高,长日照有利于M1 MDA含量的提高。G2和Q9的CAT活性在 8 h/d 处理下显著高于12、16 h/d处理(P<0.05),M1在12 h/d处理下显著高于8、16 h/d处理(P<0.05),表明短日照处理有利于G2和Q9 CAT活性的提高,日中性处理有利于M1 CAT活性的提高。M1和Q9的Sp含量在8 h/d处理下显著高于12、16 h/d处理(P<0.05),G2在16 h/d处理下显著高于8、12 h/d处理(P<0.05),表明短日照处理有利于M1和Q9 Sp的合成,长日照有利于G2 Sp的合成。G2的SOD活性在12 h/d处理下显著高于8、16 h/d处理(P<0.05),M1在8 h/d处理下显著高于12、16 h/d处理(P<0.05),Q9则在 16 h/d 处理下显著高于8、12 h/d处理(P<0.05),表明短日照处理有利于M1 SOD活性的提高,日中性处理有利于G2 SOD活性的提高,长日照有利于Q9 SOD活性的提高。G2和Q9的Pro含量在16 h/d处理下显著高于8、12 h/d处理(P<0.05),M1在8 h/d处理下显著高于12、16 h/d处理(P<0.05),表明长日照处理有利于G2和Q9 Pro含量的提高,短日照有利于M1 Pro含量的提高。
2.3 马铃薯表型性状和生理生化指标方差分析
由表3可知,光周期对马铃薯的干质量、根数有极显著影响(P<0.01),对鲜质量、总根长有显著影响(P<0.05),对株高、茎节数、茎粗、叶片数、可溶性蛋白含量、CAT活性、SOD活性、MDA含量和Pro含量无显著影响;品种对株高和根数有极显著性影响(P<0.01),对茎节数、叶片数、Pro含量的影响显著(P<0.05),对茎粗、总根长、鲜质量、干质量、可溶性蛋白含量、CAT活性、MDA含量和SOD活性无显著影响;光周期与品种互作效应对茎粗、可溶性蛋白含量、CAT活性、SOD活性、MDA含量和Pro含量有极显著影响(P<0.01),对株高、茎节数、鲜质量的影响显著(P<0.05),对干质量、根数、总根长无显著影响。
2.4 马铃薯光周期相对敏感度
光周期相对敏感度(PS)的计算公式如下:PS日中性=某日中性性状测量值/某短日照性状测量值;PS长日照=某长日照性状测量值/某短日照性状测量值。数值越大,说明该品种光周期越敏感;数值越小,说明光周期越不敏感[17]。对光周期具有显著或极显著差异的干质量、根数、鲜质量、根长和呈规律性增长的茎粗5个表型性状的相对光周期敏感度之和作为评价1个品种对光周期的敏感程度,分析3个马铃薯品种在日中性(12 h/d)和长日照(16 h/d)条件下光周期的敏感性。在日中性(12 h/d)条件下,3种供试马铃薯品种光周期相对敏感度依次为G2(1.649)>M1(1.510)>Q9(1.409),在长日照条件下,光周期相对敏感度依次为M1(1.967)>G2(1.775)>Q9(1.630),日中性条件变为长日照条件,G2、M1和Q9的光周期相对敏感值均递增,但是Q9在日中性和长日照下光周期相對敏感值比G2和M1都低。
3 討论与结论
光周期不仅可以影响植物的生长发育,同时也可以影响植物生理生化特性。有研究表明,延长光周期可以提高芽苗菜油葵的株高与叶面积[18]以及黄瓜幼苗的生长速率、茎粗与根冠比[19],适当延长光周期也可使植物进行光合作用的时间延长,从而产生更多碳水化合物支持植株生长发育。本试验中以马铃薯组培苗的干质量、株高、茎节数、根数、鲜质量、根长和茎粗为主要表型性状,研究不同光周期对不同马铃薯品种表型性状的影响,结果表明,随着光照时数的增长,G2、M1和Q9的鲜质量、干质量、茎粗以及根长均有所增加,这说明光照不仅有利于植株的生长,还能促进根系发育。这与光周期影响生菜生长的研究结果[20]一致。不同光周期对不同品种生理生化特性的影响也是不一致的。本研究发现,在短日照处理下,G2的MDA含量及CAT活性最高,M1的可溶性蛋白含量、SOD活性、Pro含量以及Q9的MDA含量、CAT活性、可溶性蛋白含量最高;在长日照处理下,G2的可溶性蛋白含量及Pro含量、M1的MDA含量以及Q9的SOD活性、Pro含量较其他光周期处理有所增加;在日中性处理下,G2的SOD活性以及M1的CAT活性也较其他光周期处理有所增加。
对作物光周期敏感性进行研究,是开展种质的适应性评价及筛选、揭示光周期敏感性遗传规律、光周期敏感相关性状QTL定位及基因克隆的基础性工作[17,21-23]。目前对马铃薯光周期敏感性鉴定的研究还未见报道,以往按照科研经验和地域性划分马铃薯光周期的敏感性,本研究选择3种不同马铃薯品种,模拟了3种日照长度,调查研究了8个表型性状和5个生理生化指标,研究模拟的日照长度差异大,适合研究马铃薯光周期敏感性。通过对干质量、根数、鲜质量和根长4个表型性状用来评价马铃薯的光周期敏感性,得出青薯(Q9)相比闽薯1号(M1)和国外2号(G2)为光周期不敏感材料。
参考文献:
[1]单建伟. 光诱导的马铃薯试管薯形成相关基因的转录组分析[D]. 武汉:华中农业大学,2013:8-9.
[2]边 纪. 世界马铃薯产业现状[J]. 新农村,2005(6):22.
[3]陈亚兰,张 健,李 定,等. 国外马铃薯种质资源的引进及利用[J]. 农业科技与信息,2013(16):26-28.
[4]陈晓仪,杨千叶,赵 琦. 不同光周期对浮萍生长及淀粉积累的影响[J]. 北方园艺,2017(22):50-54.
[5]陈 敏,李海云. 不同光周期对茄子幼苗生长的影响[J]. 北方园艺,2010(16):53-55.
[6]陈瑶瑶,严良文,刘智成,等. 苗期不同光周期处理对苦瓜性别分化及相关生理特性的影响[J]. 福建农业学报,2016,31(2):135-139.
[7]何 蔚,陈丹艳,胡晓婷,等. 不同光周期与光质配比对番茄植株生长发育的影响[J]. 西北农业学报,2018,27(4):562-570.
[8]杨晓旭,刘大军,闫志山,等. 不同光周期对菜豆生长的影响[J]. 黑龙江大学工程学报,2019,10(1):80-83.
[9]许恩婷. 金线莲开花生理与花期调控初步研究[D]. 杭州:浙江农林大学,2019:4-18.
[10]任永哲,陈彦惠,库丽霞,等. 玉米光周期反应研究简报[J]. 玉米科学,2005,13(4):86-88.
[11]贾小平,李剑峰,全建章,等. 不同光周期条件下谷子农艺性状的光周期敏感性评价[J]. 植物遗传资源学报,2018,19(5):919-924.
[12]马伟清,董道峰,陈广侠,等. 光照长度、强度及温度对试管薯诱导的影响[J]. 中国马铃薯,2010,24(5):257-262.
[13]张小川,张建虎,郭志乾,等. 光照周期对马铃薯试管薯诱导的影响[J]. 现代农业科技,2017(12):65,68.
[14]李秋琛. 水肥供应及光照对马铃薯微型薯生长发育的影响[D]. 银川:宁夏大学,2018:4-14.
[15]李鹏程,毕真真,梁文君,等. DNA甲基化参与调控马铃薯干旱胁迫响应的初步研究[J]. 作物学报,2019,45(9):1-10.
[16]邹 琦. 植物生理学实验指导书[M]. 北京:中国农出版社,2000:127-168.
[17]贾小平,全建章,王永芳,等. 不同光周期环境对谷子农艺性状的影响[J]. 作物学报,2019,45(7):1119-1127.
[18]张 欢,章丽丽,李 薇,等. 不同光周期红光对油葵芽苗菜生长和品质的影响[J]. 园艺学报,2012,39(2):297-304.
[19]李海云,韩国徽,任秋萍,等. 不同光周期对黄瓜幼苗生长的影响[J]. 西北农业学报,2009,18(3):201-203.
[20]严 妍. 温度和光周期对水培生菜生长的影响及生长模型初探[D]. 武汉:华中农业大学,2008:7-13.
[21]夏正俊. 大豆光周期反应与生育期基因研究进展[J]. 作物学报,2013,39(4):571-579.
[22]孙朝辉. 玉米光周期敏感相关性状的QTL定位与分析[D]. 郑州:河南农业大学,2009:8-13.
[23]库丽霞,孙朝辉,王翠玲,等. 玉米光周期敏感相关性状发育动态QTL定位[J]. 作物学报,2010,36(4):602-611.
