基于果实品质性状和SSR标记的李种质资源聚类
2020-03-25李雷王海燕陶尚玉陈梦微龙星雨夏惠祝进邓群仙周琼
李雷,王海燕,陶尚玉,陈梦微,龙星雨,夏惠,祝进,邓群仙*,周琼
基于果实品质性状和SSR标记的李种质资源聚类
李雷1,2,王海燕1,陶尚玉1,陈梦微1,龙星雨1,夏惠3,祝进4,邓群仙1*,周琼5
(1.四川农业大学园艺学院,四川 成都 611130;2.达州市茶果技术推广站,四川 达州 635000;3.四川农业大学果蔬研究所,四川 成都 611130;4.四川省园艺作物技术推广总站,四川 成都 611041;5.汉源县农业农村局,四川 汉源 625300)
采用25个果实性状指标和19对SSR引物对四川栽培的19份李种质资源进行主成分分析和聚类分析,并以欧式距离、前3个主成分和遗传相似性为基础,分别作系统聚类、三维排序图和UPGMA聚类。结果表明:19份李种质25个果实性状指标的平均变异系数为54.71%,其中果皮红绿色差值a值的变异系数最大,为436.14%,果核侧径的变异系数最小,为9.99%;系统聚类可将李果实分为深色系大果型和浅色系小果型;主成分分析中,前3个主成分累计贡献率为73.27%,第1主成分(果重因子)的贡献率为49.45%,第2主成分(果形与色泽因子)的贡献率为14.21%,第3主成分(营养因子)的贡献率为9.62%;以主成分PC1、PC2和PC3向量为轴的三维排序图清晰地展示出各种质在果实性状上的相对差异;SSR标记显示供试材料的遗传多样性丰富,UPGMA聚类与系统聚类和主成分三维排序的结果基本一致。
李;果实性状;微卫星标记;聚类分析

李为蔷薇科(Rosaceae)、李亚科(Prunoideae)、李属()植物[1],李果实营养丰富,是重要的夏季鲜食水果。据联合国粮食及农业组织(FAO)的统计数据,2016年,中国李收获面积和产量分别为198.57 万hm2和679.20 万t,收获面积和总产量分别占全球的75.81%和57.76%,是全球第一大生产国[2]。四川李种质资源丰富,2017年栽培面积超过7 万hm2,受生态环境和消费因素的限制,使得对李的选育带有明显的倾向性,近年来选育的‘脆蜜李’‘巴山脆李’‘羌脆李’等品种均为脆李类型,但长此以往可能会导致育成品种的遗传背景较为单一化。在长期的引种栽培和人工选择过程中,四川李种质可能发生遗传变异,对这些品种的正确鉴别和果实性状的评价,了解品种间的遗传变异和多样性,是四川李种质资源开发利用和育种改良的重要前提。
有研究发现表型差异与遗传差异之间没有必然的关系[3]。主成分和聚类分析具有直观反映作物品种资源间的遗传差异及类群性状的特点[4],在杨梅[5]、苹果[6]、核桃[7]等果树品种资源和遗传育种中被广泛使用。农艺性状评价在一定程度上容易受环境和观察者专业能力的影响,将反映表型差异的农艺性状标记与反映遗传差异的分子标记相结合能够提供更全面准确的数据[8–9]。笔者对在四川收集的19个李种质的果实大小、色差值、色素含量及营养品质等25个性状指标进行测定,同时提取叶片DNA,基于果实品质性状及SSR标记分析种质的遗传差异、类群特征和群体结构,以期为四川李种质资源鉴定评价和杂交亲本选择提供依据。
1 材料与方法
1.1 材料
19份李种质资源(表1),除茵红李采自四川省江安县桐梓镇姜庙村(105°52′E,29°46′N,海拔高度378 m)外,其余材料均采自四川省汉源县九襄镇堰坪村(102°37′~102°40′E,29°29′~29°35′N,海拔高度1 233~1 362 m)。
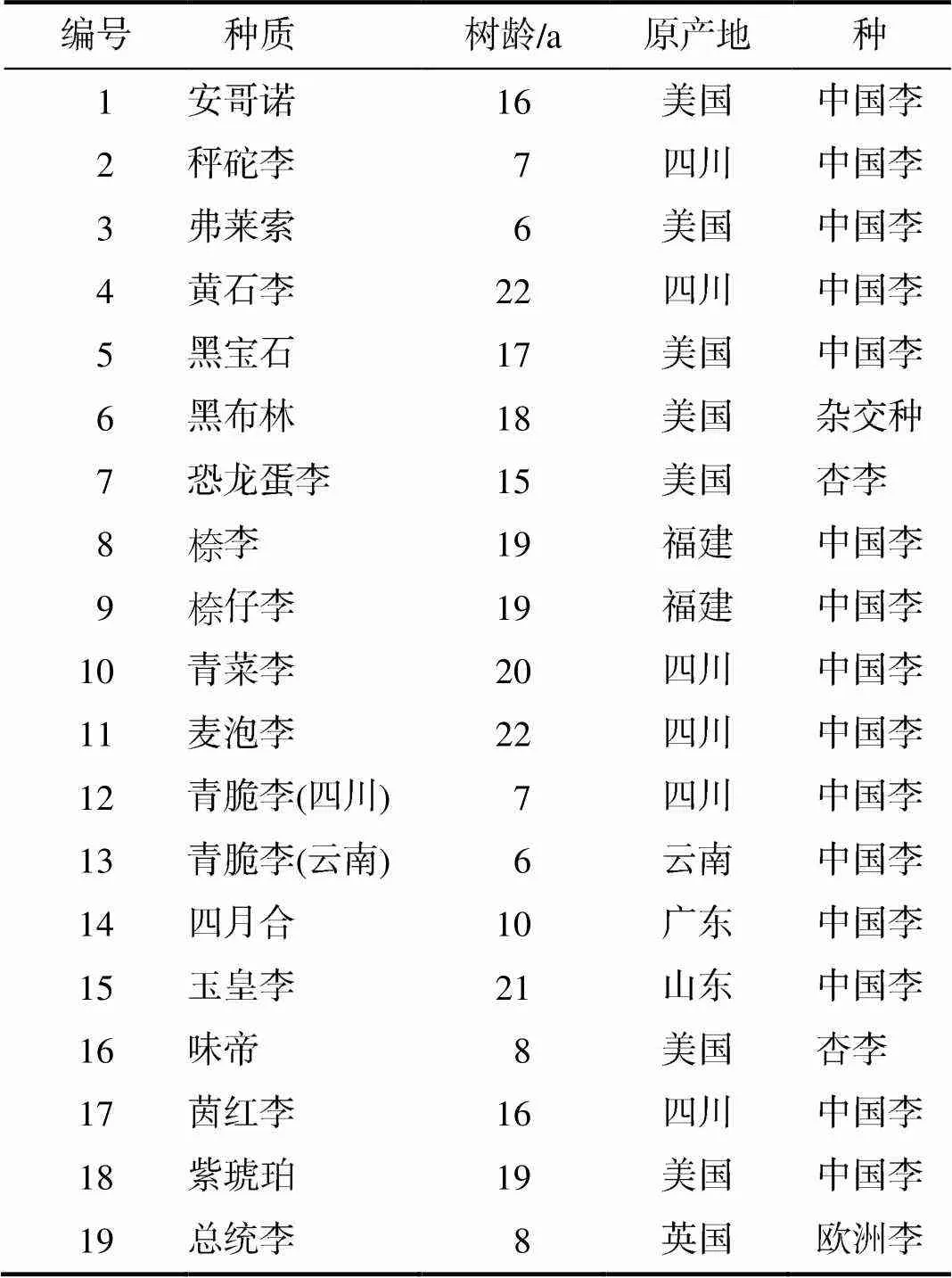
表1 李种质资源的基本信息
1.2 方法
1.2.1果实性状测定
于果实达鲜食成熟期时,每个种质选择3株树,在树体东、南、西、北4个方向树冠外围结果枝上各摘取5个着色良好、果形正常的果实,用电子天平测定果实和果核质量;用数显游标卡尺测定果实与果核的纵径、横径和侧径;用GY–4型数显硬度计测定果实硬度;用CM–2500d型分光测色计(Konica Minolta,日本)测定果实赤道部位果皮和果肉的色泽明亮度L、红绿色差值a和黄蓝色差值b。用WYT–1型手持折光仪测定李果实可溶性固形物含量;用蒽酮比色法[10]测定果肉可溶性糖含量;用氢氧化钠滴定法测定果肉可滴定酸含量;用Fe3+还原法[11]测定果肉抗坏血酸含量;用80%丙酮浸提法[10]测定果皮和果肉叶绿素与类胡萝卜素含量;用亚硝酸钠–硝酸铝比色法[12]测定果皮和果肉类黄酮含量。
1.2.2SSR标记分析
于初春采集果树新梢嫩叶,采用改良CTAB法[13]提取基因组DNA,经1%琼脂糖凝胶电泳检测合格后,使用Nanodrop one微量紫外–可见光分光光度计(Thermo Scientific,美国)测定浓度后,稀释至20 ng/μL,用于PCR扩增。从合成的87对引物中筛选出多态性高、扩增稳定的19对引物[14–19],用于SSR标记分析,用聚丙烯酰胺凝胶银然法进行条带检测。
1.3 数据处理
运用SPSS 22.0进行Duncan法方差分析;将数据进行Z–分数标准化后,采用Ward法和欧式距离进行系统聚类;使用OmicShare(http://www. omicshare.com/tools)绘制三维排序图;聚丙烯酰胺凝胶中清晰重复条带记为“1”,同一位置无条带记为“0”,构建0/1矩阵,运用POPGENE 1.32计算等位基因数、有效等位基因数、观测杂合度、期望杂合度、Shannon’s信息指数和Nei’s基因多样性;运用NTSYS 2.10进行非加权组平均法(UPGMA)聚类。
2 结果与分析
2.1 李种质的果实性状
由表2可知,李种质资源25个果实性状指标中,果皮a值变异系数最大,为436.14%,果核侧径的变异系数最小,仅为9.99%;果皮叶绿素和类胡萝卜素分别是色素含量指标中变异系数最大和最小的,分别为71.62%和33.01%;抗坏血酸和可溶性固形物分别是果实营养品质中变异系数最大和最小的,分别为61.20%和12.43%。整体来看,不同李种质果实的可溶性固形物含量、可溶性糖含量、果肉L值、果实大小(纵径、横径和侧径)和果核大小(纵径、横径和侧径)的差异较小,变异系数低于20%。种质间抗坏血酸含量、果皮a值、果皮b值、果肉a值、果皮叶绿素含量、果皮类黄酮含量和果肉叶绿素含量的差异较大,变异系数高于50%。从图1可以看出,不同李种质间果实色泽差异较大。不同李种质果实性状的差异主要是色差值与色素含量,果皮、果肉色差(*、*和*)和色素含量(叶绿素、类胡萝卜素和类黄酮)的平均变异系数为178.05%、62.42%、51.74%和50.23%。值得注意的是,果皮和果肉值变异系数均高于L值的,*值的最小,表明果实的主要色差为红绿色差。

图1 李果实成熟期的色泽
2.2 基于果实性状的系统聚类
基于李果实25个性状指标,对19份李种质资源进行系统聚类,结果如图2所示。李种质可分为3个类群。G1类群包含8份种质(秤砣李、黄石李、青菜李、麦泡李、青脆李(四川)、青脆李(云南)、四月合和茵红李),G1类群的果皮b值、果肉b值、果皮及果肉叶绿素含量均显著高于G2和G3类群,而果皮a值、果皮b值、单果质量、果实纵径和果核质量均显著低于G2和G3类群(表3)。G1类群中,除茵红李果皮有部分区域呈红色外,其他种质均为浅色系(黄绿色),小果型。G2类群包含玉皇李、味帝和紫琥珀3个种质,果肉L值、果肉b值和硬度均显著低于G1和G3类群的。G3类群包含安哥诺弗莱索、黑宝石、黑布林、恐龙蛋、㮈李、㮈仔李和总统李等8个种质,其果核质量、果核侧径和果皮类黄酮含量均显著高于G1和G2类群,除㮈李和㮈仔李外,另外6个种质均为深色系(紫黑色),大果型。

图2 李种质资源的系统聚类

表3 不同类群的李种质果实性状测定值
同一行数据不同小写字母表示差异显著(0.05)。
2.3 基于果实性状主成分的三维排序
李果实性状主成分分析的特征向量值列于表4。前5个主成分的方差贡献率分别为49.45%、14.21%、9.62%、7.77%和4.95%,累计方差贡献率为86.01%。第一主成分(PC1)的特征值为12.36,主要与单果质量和果实侧径相关,因此可以将第一主成分称为果重因子。第二主成分(PC2)的特征值为3.55,主要与果肉L值、果实横径和果肉b值相关,可以称为果形与色泽因子。第三主成分(PC3)的特征值为2.40,主要与可溶性固形物含量和可溶性糖含量相关,可以称为营养因子。第四主成分(PC4)的特征值为1.94,主要与抗坏血酸含量和果肉a值相关。第五主成分(PC5)的特征值为1.24,主要与可溶性糖含量和硬度相关,其中可溶性糖含量为负向因子。

表4 李果实性状主成分分析的特征向量值

表4(续)
利用前3个主成分绘制19份李种质的三维排序图(图3),PC1轴0点左侧的群体较右侧的群体更为分散,PC1轴上差异最大的是安哥诺与青菜李,PC2轴上差异最大的是安哥诺与味帝,PC3轴上差异最大的是㮈李与紫琥珀。
2.4 基于SSR标记的李种质资源多样性和聚类
所选的19对引物均能扩增出清晰整齐、多态性高的条带,图4为引物CPSCT034在19份李种质中的扩增结果。从表5可以看出,19对引物在19份种质中扩增出的等位基因数最大值为9 (UDP96–010),最小值为3(CPSCT044),平均等位基因数为5.06;有效等位基因数最大值为6.45 (UDP96–010),有效等位基因数平均值为3.43。Shannon’s信息指数的最大值为2.00(UDP96–010),平均值为1.30。Nei’s基因多样性的最大值为0.84(UDP96–010),平均值为0.66。表明供试李种质的遗传变异度较高,遗传多样性丰富。
基于SSR标记的聚类如图5所示,19份李种质被分为3组,第1组(K1)包含6份种质,其中安哥诺、弗莱索、黑宝石、黑布林、紫琥珀来自美国,总统李来自英国。第2组(K2)包含5份种质(恐龙蛋李、㮈李、㮈仔李、玉皇李、味帝),K21亚组包括2个来自福建的种质(㮈李、㮈仔李)和2个美国种质(恐龙蛋李、味帝);玉皇李独立成为K22亚组,这与玉皇李在G2类群中的聚类模式基本一致。第3组(K3)包含8个种质(秤砣李、黄石李、青菜李、麦泡李、青脆李(四川)、青脆李(云南)、四月合、茵红李),与G1类群的8个种质相同;K31亚组包含四川和云南的7个种质,来自广东的四月合独立成为K32亚组。
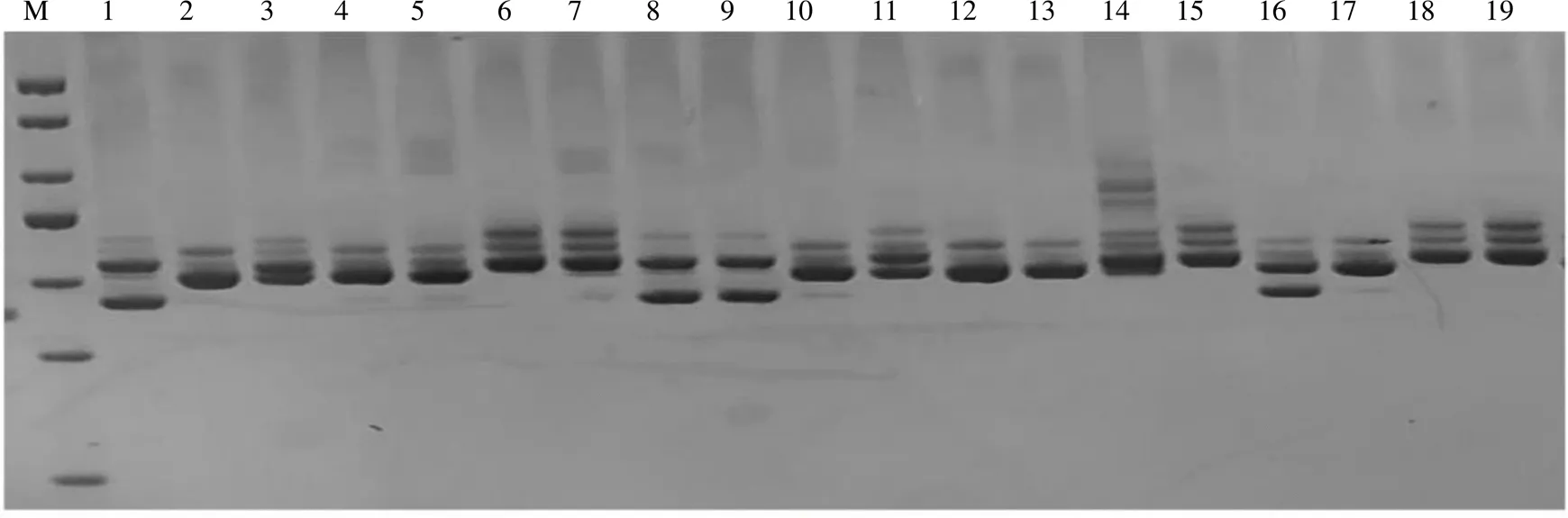
M为DNA DL500 Maker;1~19为种质编号。
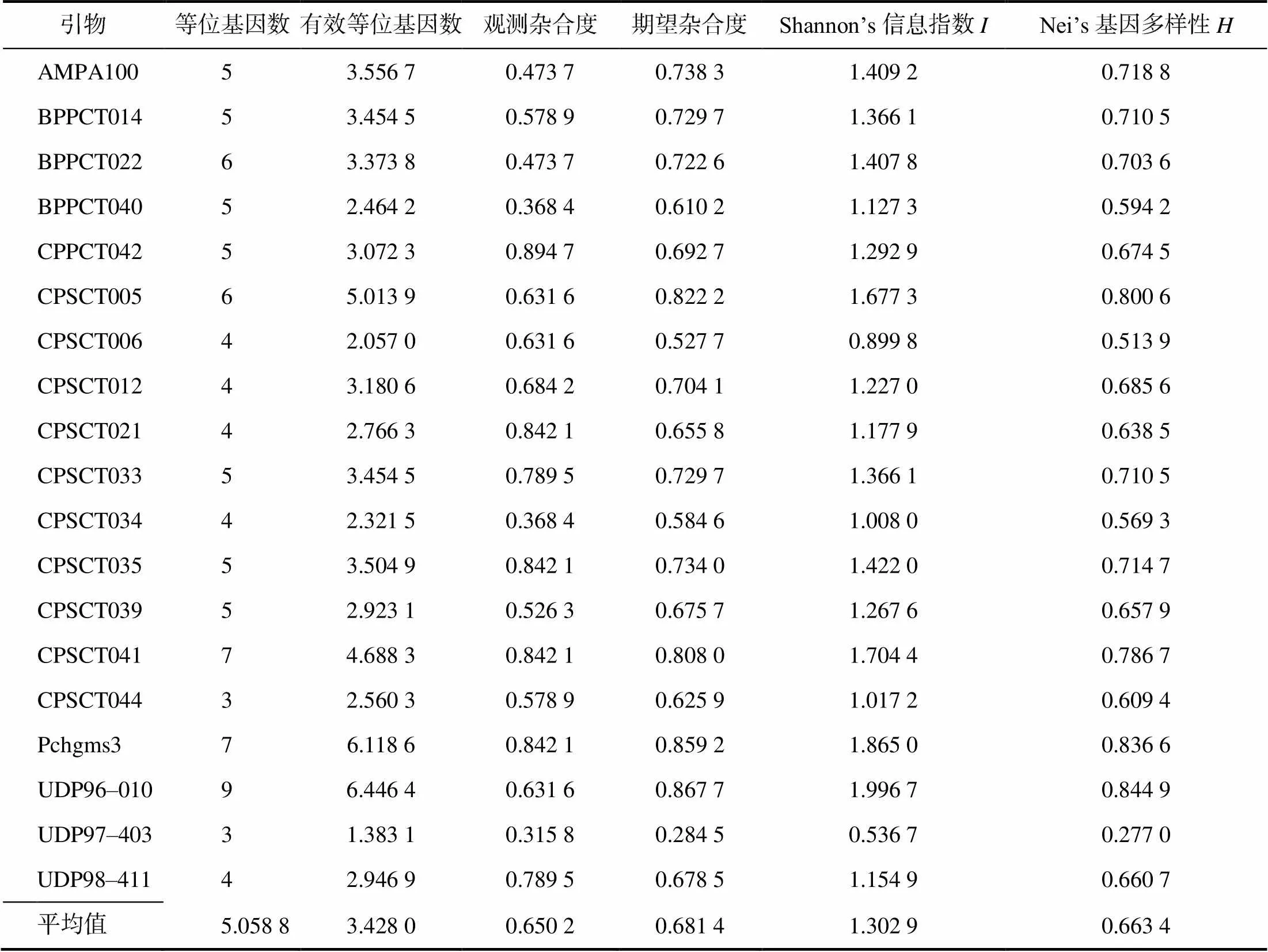
表5 19份李种质资源在19对引物中的遗传多样性指数

图5 李种质资源基于SSR标记的UPGMA聚类
3 结论与讨论
19份李种质资源的25个果实性状指标的变异系数为9.99%~436.14%,平均值为54.71%,说明种质间果实性状差异极大,变异范围较广,多样性丰富,这与前人在欧洲李[20]和中国李[21]中的研究结果基本一致。果实核型大小的变异度最小,与王美军等[22]在刺葡萄中的研究结果一致。李果实色泽类型极为丰富,果皮彩色分6种类型,果肉色泽分8种类型[1]。果实色泽多被当作描述型指标,缺少定量数据。本试验测定了果实色差和主要色素物质含量,为李果实色泽评价提供了量化数据。果皮和果肉a值的变异系数分别为436.14%和153.80%,远高于其他指标,除去果皮和果肉a值后感官指标的平均变异系数仅为28.88%,此时平均变异最丰富的是色素含量指标,这与果实a值变异系数较大相符合。本试验中的李种质果实涵盖了大部分的色泽类型,这可能是李种质间果实a值差异较大的主要原因。
19份李种质的遗传多样性参数平均值低于孙萍等[23]对47份李种质的研究结果,高于左力辉等[24]对24个不同产地中国李品种中的分析结果和孙莉莉等[25]在寒地李和杏中的研究结果,表明供试李种质具有中等至较高水平的遗传多样性。
与果实单果质量和大小相关的性状指标被确定为第一主成分(PC1),方差贡献率高达49.45%,表明单果质量对李果实的评价意义重大,这与前人对枣[26]、猕猴桃[27]和欧洲李[20]果实因子的评价基本一致,但与对苹果[6]的研究差异较大,这可能与果树种类及育种者对不同树种果实性状的关注度不同有关。大果、丰产是李重要的育种目标,与果质量、果形密切相关的指标被提取成第一主成分因子,表明供试种质存在较大的遗传改良潜力。试验中,原产中国南方的李种质果实普遍偏小,国外李种质大果型较多,这与郁香荷等[21]的研究结果相似。根据刘文东[28]对李杂交子代遗传规律的分析,小果型亲本对后代果型影响极大,不宜作为选育大果型品种的亲本;浅色系品种与深色系品种杂交,后代多表现为深色系,这种遗传倾向在杂交亲本选配时应重点考虑。此外,以国外品种为主的类群在单果质量、果实大小和类黄酮含量上均高于国内品种类群,表明引种的国外品种可作为国内李品种改良的材料。
本研究中,基于果实性状的系统聚类和基于SSR标记的UPGMA聚类呈现出相似的聚类模式,即大部分国内品种、国外品种分别聚为一类,系统聚类中的G1类群和UPGMA聚类中K3组中的材料完全相同,8份材料均来自中国南方。以PC1、PC2和PC3为坐标轴作三维排序,结果与系统聚类和UPGMA聚类基本一致,但更为直观、形象。尤其是三维排序图揭示了各品种在单果重、果形和色泽等方面的相对差异。三维排序图显示来自南方的李品种在空间分布较为集中,来自国外的李品种分布更为分散,这在一定程度上说明了南方李品种更为原始,国外李品种则有着更为丰富的多样性[29]。
[1] 张加延,周恩.中国果树志·李卷[M].北京:中国林业出版社,1998:13–26.ZHANG J Y,ZHOU E.China Fruit-Plant Monographs·Plum Flora[M].Beijing:China Forestry Press,1998:13–26.
[2] Food and Agriculture Organization of the United Nations. Food and agriculture organization of the united nations fao-sate database collections[DB/OL].(2019–01–18) [2019–02–21].http://www.fao.org/faostat/zh/?#data/QC.
[3] 卓书斌,黄小凤,袁志永,等.台湾青枣不同品种表型差异和遗传多样性研究[J].现代农业科学,2009,16(2):13–16.ZHUO S B,HUANG X F,YUAN Z Y,et al.Study on the phenotypic difference and genetic diversity of different cultivars of Taiwan green jujube[J].Modern Agricultural Sciences,2009,16(2):13–16.
[4] 赵建栋,李秀莲,史兴海,等.苦荞品种(系)聚类分析[J].农学学报,2016,6(8):12–17.ZHAO J D,LI X L,SHI X H,et al.Cluster analysis ofvarieties(lines)[J].Journal of Agriculture,2016,6(8):12–17.
[5] 王伟,吕旭健,张玉,等.基于聚类分析和主成分分析法的杨梅营养品质评价研究[J].食品工业科技,2017,38(1):278–280.WANG W,LVU X J,ZHANG Y,et al.Evaluation of nutritional quality of red bayberry based on cluster analysis and principal component[J].Science and Technology of Food Industry,2017,38(1):278–280.
[6] 徐吉花,赵政阳,王雷存,等.苹果果实品质评价因子的选择研究[J].干旱地区农业研究,2011,29(6):269–274.XU J H,ZHAO Z Y,WANG L C,et al.Selection of factors for apple fruit quality evaluation[J].Agricultural Research in the Arid Areas,2011,29(6):269–274.
[7] 蒲光兰,肖千文,吴开志,等.四川核桃种质资源表型多样性研究[J].湖南农业大学学报(自然科学版),2014,40(2):162–167. PU G L,XIAO Q W,WU K Z,et al.Research on the phenotypic diversity of walnut germplasm esources in Sichuan[J].Journal of Hunan Agricultural University (Natural Sciences),2014,40(2):162–167.
[8] 余智城,何雪娇,曹明华.基于表型性状和SSR标记的20份香蕉材料遗传多样性研究[J].中国南方果树,2014,43(3):10–14. YU Z C,HE X J,CAO M H.Genetic diversity of 20 banana germplasm based on phenothpic characteristics and ssr markers[J].South China Fruits,2014,43(3):10–14.
[9] 王姣,刘崇怀,樊秀彩,等.河南境内桑叶葡萄种内形态和遗传多样性分析[J].果树学报,2008,25(4):496–500.WANG J,LIU C H,FAN X C,et al.Study on the biodiversity of the morphology and polymorphism based on SSR markers for Vitis ficifolia native to Henan province in China[J].Journal of Fruit Science,2008,25(4):496–500.
[10] 王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006:134–136,202–204.WANG X K.Experimental Principles and Techniques of Plant Physiology and Biochemistry[M].Beijing:Higher Education Press,2006:134–136,202–204.
[11] KAMPFENKEL K,VAN MONTAGU M,INZE D. Effects of iron excess onplants (implications to oxidative stress)[J].Plant Physiology,1995,107(3):725–735.
[12] 唐巧玉,周毅峰,朱玉昌,等.金橘皮中黄酮类物质的提取及其体外抗氧化活性研究[J].农业工程学报,2008,24(6):258–261.TANG Q Y,ZHOU Y F,ZHU Y C,et al.Extraction of flavone compounds frompeel and its antioxidation in vitro[J].Transactions of the CSAE,2008,24(6):258–261.
[13] 张慧蓉,乔玉山,曹尚银,等.几个杏李品种成熟叶片基因组DNA的提取[J].江西农业学报,2008,20(10):4–6. ZHANG H R,QIAO Y S,CAO S Y,et al.Extraction of genomic DNA from mature leaves of some varieties of[J].Acta Agriculturae Jiangxi,2008,20(10):4–6.
[14] HAGEN L S,CHAI B J ,FADY B,et al.Genomic and cDNA microsatellites from apricot (L.)[J].Molecular Ecology Notes,2010,4(4):742–745.
[15] DIRLEWANGER E,COSSON P,TAVAUD M,et al. Development of microsatellite markers in peach [(L.) Batsch] and their use in genetic diversity analysis in peach and sweet cherry(L.)[J].Theoretical and Applied Genetics,2002,105(1):127–138.
[16] ARANZANA M J,GARCIA-MAS J,CARBOJ,et al. Development and variability analysis of microsatellite markers in peach[J].Plant Breeding,2002,121:87–92.
[17] MNEJJA M,GARCIA-MAS J,HOWAD W,et al. Simple-sequence repeat (SSR) markers of Japanese plum (Lindl.) are highly polymorphic and transferable to peach and almond[J].Molecular Ecology Notes,2004,4(2):163–166.
[18] SOSINSKI B,GANNAVARAPU M,HAGER L D,et al. Characterization of microsatellite markers in peach [(L.) Batsch][J].Theoretical and Applied Genetics,2000,101(3):421–428.
[19] TESTOLIN R,MARRAZZO T,CIPRIANI G,et al.Microsatellite DNA in peach (L.Batsch) and its use in fingerprinting and testing the genetic origin of cultivars[J].Genome,2000,43(3):512–520.
[20] 孙琪,廖康,耿文娟,等.新疆欧洲李资源枝叶形态及果实感官特征差异分析[J].新疆农业大学学报,2015,38(1):18–24. SUN Q,LIAO K,GENG W J,et al.Characteristic variance analysis on morphology of branches-leaves and fruit sense-organ ofL.resources in Xinjiang[J].Journal of Xinjiang Agricultural University,2015,38(1):18–24.
[21] 郁香荷,章秋平,刘威生,等.中国李种质资源形态性状和农艺性状的遗传多样性分析[J].植物遗传资源学报,2011,12(3):402–407. YU X H,ZHANG Q P,LIU W S,et al.Genetic diversity analysis of morphological and agronomic characters of chinese plum (Lindl.) germplasm[J]. Journal of Plant Genetic Resources,2011,12(3):402– 407.
[22] 王美军,黄乐,刘昆玉,等.刺葡萄叶与花和果实及种子的表型性状研究[J].湖南农业大学学报(自然科学版),2016,42(5):489–495. WANG M J,HUANG L,LIU K Y,et al.Phenotypic traits of leaf, flower, fruit and seed of[J].Journal of Hunan Agricultural University(Natural Sciences),2016,42(5):489–495.
[23] 孙萍,林贤锐,沈建生.基于SSR标记的李种质资源遗传多样性研究[J].江西农业学报,2017,29(2):33–39. SUN P,LIN X R,SHEN J S.Study on genetic diversity of plum germplasm resources based on SSR markers[J]. Acta Agriculturae Jiangxi,2017,29(2):33–39.
[24] 左力辉,韩志校,梁海永,等.不同产地中国李资源遗传多样性SSR分析[J].园艺学报,2015,42(1):111–118. ZUO L H,HAN Z J,LIANG H Y,et al.Analysis of genetic diversity offrom different producing areas by SSR markers[J].Acta Horticulturea Sinica,2015,42(1):111–118.
[25] 孙莉莉,牟蕴慧,甄灿福,等.寒地李和杏遗传多样性SSR分析[J].北方园艺,2017(13):50–55.SUN L L,MU Y H,ZHEN C F,et al.Genetic diversity of SSR analysis plum and apricot in cold region[J]. Northern Horticulture,2017(13):50–55.
[26] 刘隋赟昊,韩超,于婷,等.引进新疆的48个枣品种果实与果核性状分析[J].经济林研究,2016,34(4):37–46. LIU S Y H,HAN C,YU T,et al.Analysis of fruit and stone characters in 48cultivars introduced in Xinjiang[J].Nonwood Forest Research,2016,34(4):37–46.
[27] 秦红艳,许培磊,艾军,等.软枣猕猴桃种质资源果实品质、表型性状多样性及主成分分析[J].中国农学通报,2015,31(1):160–165.QIN H Y,XU P L,AI J,et al.Diversity of fruit quality and phenotypic traits ofplanch germplasm resources and their principal component analysis[J].Chinese Agricultural Science Bulletin,2015,31(1):160–165.
[28] 刘文东.李树杂交后代亲子性状遗传变异规律[J].中国林副特产,2013(5):96–97. LIU W D.Genetic variation of plum tree hybrid offspring[J].Forest By-Product and Speciality in China,2013(5):96–97.
[29] LIU W S,LIU D C,FENG C J,et al.Genetic diversity and phylogenetic relationships in plum germplasm resources revealed by RAPD markers[J].The Journal of Horticultural Science and Biotechnology,2006,81(2):242–250.
Cluster analysis of plum germplasm resources based on fruit quality traits and SSR markers
LI Lei1,2, WANG Haiyan1, TAO Shangyu1, CHEN Mengwei1, LONG Xingyu1, XIA Hui3, ZHU Jin4, DENG Qunxian1*, ZHOU Qiong5
(1.College of Horticulture, Sichuan Agricultural University, Chengdu, Sichuan 611130, China; 2.Tea and Furit Technology Extension Station of Dazhou, Dazhou, Sichuan 635000, China; 3.Institute of Pomology and Olericulture, Sichuan Agricultural University, Chengdu, Sichuan 611130, China; 4.Sichuan Horticultural Crop Technology Extension Station, Chengdu, Sichuan 611041, China; 5.Hanyuan County Bureau of Agriculture and Rural Affairs, Hanyuan, Sichuan 625300, China)
Principal component analysis andcluster analysis of19 plum germplasm resources which planted in Sichuan were conducted using 25 fruit characters and 19 SSR markers. The plum germplasm were classified based on Euclidean distance, the first three principal components and genetic similarity, as well as systematic cluster, three-dimensional ordination plot and UPGMA clustering analysis. Results showed that the average coefficient of variation of the 25 fruit characters was 54.71%, the variation coefficient ofvalue of the peel was the highest (436.14%) and the variation coefficient of stone lateral dinmeter was the lowest (9.99%). The system clustering mainly divided these varieties into large fruit with deep color, small fruit with light color. In the principal component analysis, the first three principal components account for 73.27% of variation among the varieties, the first principal component (PC1) which regarded as fruit weight factor contributed 49.45%, the second principal component (PC2) as fruit shape and the color factor contributed 14.21%, and the third principal component (PC3) as nutritional factor contributed 9.62%. The three-dimensional ordination plot with PC1, PC2 and PC3 vectors clearly showed the relative differences of the main characters for the 19 plum germplasm resources. SSR markers showed that the genetic diversity of the tested materials was rich. The results of UPGMA clustering were similar to those of systematic clustering and three-dimensional ordination plot.
plum; fruit characters; simple sequence repeats makers; cluster analysis
10.13,331/j.cnki.jhau.2020.01.007
S662.302.3
A
1007-1032(2020)01-0038-10
2019–04–19
2019–12–06
四川省科技支撑项目(2013FZ0036);四川农业大学“双支计划”基金项目(03572783);四川农业大学与四川省汶川县校地合作项目(1922339018)
李雷(1993—),男,四川平昌人,硕士研究生,主要从事果树栽培与技术研究,sicaulilei@foxmail.com;
,邓群仙,博士,教授,主要从事果树栽培理论与技术研究,1324856299@qq.com
李雷,王海燕,陶尚玉,陈梦微,龙星雨,夏惠,祝进,邓群仙,周琼.基于果实品质性状和SSR标记的李种质资源聚类[J].湖南农业大学学报(自然科学版),2020,46(1):38–47.
LI Lei, WANG H Y, TAO S Y, CHEN M W, LONG X Y, XIA H, ZHU J, DENG Q X, ZHOU Q. Cluster analysis of plum germplasm resources based on fruit quality traits and SSR markers[J].Journal of Hunan Agricultural University(Natural Sciences), 2020, 46(1):38–47.
http://xb.hunau.edu.cn
责任编辑:罗慧敏
英文编辑:罗维
