黑棘鲷仔鱼饥饿实验及不可逆点的确定*
2020-03-25杨育凯林黑着王志勇谢仰杰郭奕惠黄小林
杨育凯 林黑着 王志勇 谢仰杰 郭奕惠 黄小林 李 涛
黑棘鲷仔鱼饥饿实验及不可逆点的确定*
杨育凯1林黑着1王志勇2①谢仰杰2郭奕惠1黄小林1李 涛1
(1. 中国水产科学研究院南海水产研究所 农业农村部南海渔业资源开发利用重点实验室 广州 510300;2. 集美大学水产学院 农业农村部东海海水健康养殖重点实验室 厦门 361021)
在水温18℃~19℃条件下,采用实验生态学方法研究了饥饿对黑棘鲷()仔鱼存活、生长发育及行为学特征的影响。通过测定仔鱼的初次摄食率和饥饿不可逆点(PNR),确定了初次投喂的最佳时间。结果显示,黑棘鲷仔鱼3日龄开口摄食,4日龄卵黄囊消失,6日龄油球消失,混合营养期仅3 d,属容易遭受饥饿胁迫的鱼类。仔鱼开口时初次摄食率仅为30%,之后迅速提高,至5日龄达到最高的90%,之后逐渐下降,7日龄降至45%,到达PNR。饥饿组仔鱼在6日龄后开始出现生理性萎缩,全长与对照组相比差异显著(<0.05),进入PNR后,仔鱼活动能力显著下降,身体出现扭曲、畸形,死亡率急剧升高,至10日龄全部死亡。黑棘鲷仔鱼耐受饥饿能力较弱,建议开始投饵的最适时机为仔鱼开口后的4 d内。
黑棘鲷;仔鱼;饥饿;不可逆点
黑棘鲷(),又称黑鲷,俗名黑加吉、黑立等,隶属鲈形目(Perciformes)、鲷科(Sparidae)、棘鲷属(),广泛分布于我国、朝鲜半岛和日本沿海(吴仁协等, 2011)。黑棘鲷生长迅速、食性广泛、适应性强,肉质鲜美、营养丰富、经济价值高,是我国海水增养殖的优良品种和重要的捕捞对象。
引起仔鱼死亡的因素有很多,其中,饥饿是主要因素之一(殷名称, 1991)。仔鱼孵化后,要经历从内源性营养到外源性营养转变的关键时期,若不能持续获得食物,经过一段时间将彻底丧失摄食能力,达到饥饿不可逆点(Point of no return, PNR),最终导致死亡。PNR即初次摄食期仔鱼耐受饥饿的时间临界点,最早是由Blaxter等(1963)提出的,是从生态学角度测定仔鱼饥饿耐受能力的一项重要指标。利用饥饿实验确定仔鱼初次摄食率和PNR,对于研究鱼类早期生活史及苗种培育工作有着十分重要的意义。近年来,国内相关研究已在多种鱼类中开展(陈淑吟等, 2018; 段国庆等, 2018; 高小强等, 2015; 许晓娟等, 2010),但有关黑棘鲷的研究尚未见报道。作者就饥饿对黑棘鲷仔鱼内源性营养的吸收及生长发育的影响进行研究,并测定仔鱼的初次摄食率和饥饿PNR,旨在全面了解黑棘鲷早期发育过程中的摄食特点和饥饿耐受能力,为大规模培苗提供参考。
1 材料与方法
1.1 材料
实验于2016年3月在中国水产科学研究院南海水产研究所深圳实验基地进行。黑棘鲷初孵仔鱼来自深圳大亚湾东山渔排养殖2龄以上亲鱼,通过肌肉注射促黄体素释放激素A3 (LHRH-A3)进行人工催产,自然产卵孵化获得。
1.2 方法
1.2.1 仔鱼培育 仔鱼孵化后随机分为A、B和C 3组,每组2000尾放入80 L圆形塑料桶中,微充气,每天吸底,换水1次,换水量为1/3。A组作为正常对照组,从仔鱼开口前1 d开始,每天定时投喂经小球藻()和强化的褶皱臂尾轮虫(),密度为10~15个/ml;B和C组设为饥饿组,不投饵。实验用海水经三级过滤,自然水温为18℃~19℃,盐度为30~31,溶氧(DO)≥5.0 mg/L,pH为7.8~8.0。
1.2.2 饥饿实验 实验开始后,每天检查记录各组死亡情况,观察仔鱼形态发育和行为特征。定时从A和B组中各取10尾仔鱼,在带有目测微尺的显微镜下测量全长、卵黄囊长径、短径和油球直径,以全长–日龄关系曲线和特定生长率(SGR)比较饥饿对仔鱼生长的影响,并计算全长的组内差异系数(CV)。
卵黄囊体积=4/3×π×(/2)2×/2
式中,为卵黄囊短径,为卵黄囊长径。
油球体积=4/3×π×(/2)3
式中,为油球直径。
SGR(ln2-ln1)/(2-1)×100%
式中,2、1分别是实验结束时和实验开始时仔鱼的全长,2、1分别表示实验仔鱼日龄。
1.2.3 初次摄食率和PNR 黑棘鲷仔鱼开口后,每天从C组随机取出20尾仔鱼,放入500 ml烧杯中。微充气并投喂轮虫,密度为10~15个/ml。4 h后,将仔鱼取出,5%福尔马林固定,在显微镜下,解剖、观察仔鱼摄食情况,记录摄食的仔鱼数量,并计算初次摄食率。
初次摄食率=摄食仔鱼数/样本仔鱼数×100%
PNR以黑棘鲷孵化后的日龄表示,每日测定饥饿组仔鱼的初次摄食率,当所测得的仔鱼初次摄食率低于最高摄食率的50%时,即为PNR的时间(Blaxter, 1963)。
1.3 数据分析
实验所得数据用Excel 2003软件处理和绘图,并运用SPSS 19.0软件进行检验法统计分析。
2 结果
2.1 黑棘鲷仔鱼卵黄囊和油球的吸收
黑棘鲷受精卵卵径为(0.89±0.01) mm,体积为(0.3717±0.0174) mm3(=10),在水温为18℃~19℃条件下,约36 h孵化出膜。初孵仔鱼卵黄囊长径为(1.03± 0.06) mm,短径为(0.61±0.02) mm,体积为(0.1997± 0.0176) mm3;油球直径为(0.21±0.01) mm,体积为(0.0052±0.0007) mm3(表1)。仔鱼内源性的吸收顺序为先卵黄囊后油球,孵化后,卵黄囊吸收迅速,第1天即消耗总体积的70%,至4日龄完全吸收;油球吸收速度相对较慢,第1天消耗总体积的30%;3日龄前,2组仔鱼油球消耗差异不大,之后,饥饿组油球吸收速度放缓,与对照组开始出现显著差异(<0.05);至6日龄,2组油球完全吸收,仔鱼进入外源性营养阶段。
2.2 黑棘鲷仔鱼初次摄食率和PNR
在培育水温为18℃~19℃条件下,经过不同饥饿天数后,黑棘鲷仔鱼首次投饵的初次摄食率变化水平见图1。在开口当天(3日龄),投饵4 h后,仔鱼初始摄食率为30%;之后,迅速提升,5日龄饥饿仔鱼初始摄食率达到最高的90%;随后逐渐下降,7日龄降至45%,为最高初次摄食率的1/2,仔鱼耐受饥饿的PNR即为7日龄。8日龄进行首次投饵,仔鱼初始摄食率仍有约15%,但此时的仔鱼已错过了正常构建外源营养吸收代谢关系的时机,无法再恢复到正常摄食水平;至9日龄,饥饿仔鱼不再摄食,已死亡殆尽。由此可见,饥饿仔鱼具摄食能力时间为孵化后的第3~6天,黑棘鲷仔鱼最适的起始投饵时间为仔鱼开口后的4 d内。
2.3 饥饿对黑棘鲷仔鱼生长和存活的影响
饥饿组和对照组仔鱼全长生长情况见表2和图2。3日龄前,随着内源性营养的快速吸收,2组仔鱼均保持较快的生长速度,全长增长明显;之后,生长速度逐渐放缓,5日龄前2组全长差异不显著(>0.05)。5日龄后,随着卵黄囊和油球的完全吸收,仔鱼进入外源性营养阶段,2组仔鱼生长速度开始出现分化。对照组通过摄食和吸收外界食物营养进入平稳生长阶段,至9日龄,平均全长达3.74 mm,最大全长超过4 mm,显著高于同日龄的饥饿组仔鱼(<0.05),摄食仔鱼全长总体呈对数增长,全长()与日龄()符合关系式:=0.6491ln+2.2215 (2=0.9594);饥饿组仔鱼在6日龄后出现生长停滞和小幅萎缩现象,至9日龄平均全长仅为3.14 mm,全长与日龄符合多项式=–0.03062+0.4166+1.9325 (2=0.8786)。2组仔鱼的特定生长率(SGR)也有明显差异,对照组9日龄仔鱼SGR为6.41%,明显高于饥饿组仔鱼的4.47%。
表1 黑棘鲷仔鱼卵黄囊和油球的吸收(=10)

Tab.1 Characteristics of yolk sac and oil globule in A. schlegelii larvae (n=10)
注:同日龄同指标的不同上标字母表示组间差异显著(<0.05),下同
Note: Data in the same age and same index with different superscripts are significantly different (<0.05). The same as below

图1 黑棘鲷仔鱼初次摄食率
黑棘鲷仔鱼存活情况见图3。2组仔鱼在开口前死亡率均较低(约15%),前期死亡率差异不大,5日龄对照组死亡率为32.5%,饥饿组为30.4%。6日龄后对照组仔鱼过渡到外源营养期,死亡率明显降低,7日龄的死亡率为49%,至10日龄累计死亡率为55.2%;而饥饿组仔鱼因无法获取外界营养而加剧死亡,至7日龄即死亡过半(57.3%),10日龄全部死亡。
表2 饥饿对黑棘鲷仔鱼生长的影响(=10)

Tab.2 Effect of growth on A. schlegelii larvae (n=10)
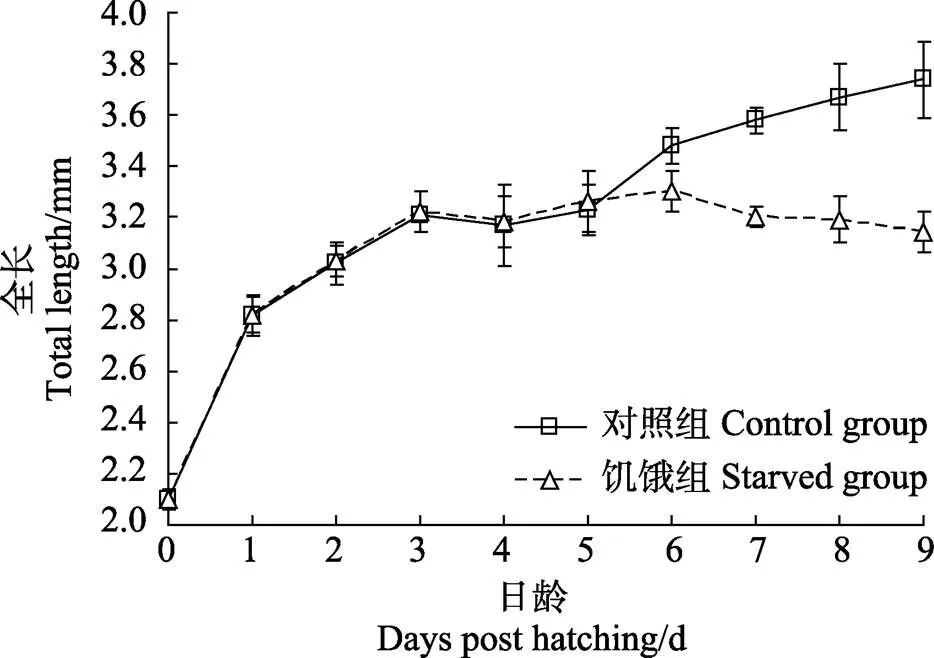
图2 饥饿对黑棘鲷仔鱼生长的影响

图3 饥饿对黑棘鲷仔鱼存活的影响
2.4 饥饿对黑棘鲷仔鱼形态发育和行为学特征的影响
在水温为18℃~19℃条件下,黑棘鲷仔鱼形态发育特征如图4所示。初孵仔鱼全长为(2.10±0.04) mm (=10),鱼体透明,身体前半部包在卵黄囊中,头部紧贴卵黄囊前端而下倾,两侧有2个圆形眼囊,未见色素,卵黄囊呈卵圆形,长径约占仔鱼全长的1/2,油球扁圆形,位于卵黄囊后端,仔鱼头部、背部和尾部有黑色素分布,肠管细线状,紧贴于卵黄囊上方,口和肛门为封闭状态(图4.1);仔鱼漂浮在水面上,游泳能力较弱,可短暂窜游,无趋光与集群现象。3日龄仔鱼身体细长,卵黄囊消失殆尽,眼球布满黑色素,视觉形成,尾部和消化道腹面色素明显,口已开启,下颌突出,开始摄食,出现胸鳍鳍褶,胃出现,消化道变粗,肛门开启(图4.2);仔鱼开始出现巡游式运动,具有集群与趋光性。4日龄仔鱼卵黄囊完全消失,油球继续吸收,尾部色素面积减少,胃、肠分化明显,胃部变大,胸鳍呈叶片状(图4.3);仔鱼运动能力增强。

图4 饥饿对黑棘鲷仔鱼形态发育的影响
1:初孵仔鱼;2~3分别为3、4日龄仔鱼;4~6分别为对照组6、8、9日龄仔鱼;7~9分别为饥饿组6、8、9日龄仔鱼
1:Newly hatched larvae; 2 and 3 are 3and 4days post hatching larvae respectively; 4, 5 and 6 are control group larvae of 6, 8, and 9days post hatching respectively; 7, 8, and 9 are starved group larvae of 6, 8, and 9days post hatching respectively
黑棘鲷对照组和饥饿组仔鱼在4日龄后形态和行为特征开始出现较大差异,对照组仔鱼游动能力进一步增强,大量集群巡游觅食,受惊吓后游动迅速分散;饥饿组仔鱼生长减缓,游动能力变弱,对外界刺激反应迟钝,约50%出现离群现象。至6日龄,仔鱼油球吸收完全,进入外源性营养阶段,此时,对照组仔鱼高度集群,胃部膨胀,充满食物,肠径变粗,胸角出现,胸鳍变大,脊节明显,仔鱼全长增加明显 (图4.4);饥饿组仔鱼身体瘦弱,胸角突出(图4.7),仔鱼生长停滞,静止或四处狂游,有些先头下尾上倒挂于水体中上层,后漂浮侧卧于水面,受惊扰后作螺旋状下游,不再成群游动,摄食能力较弱。8和9日龄对照组仔鱼摄食能力进一步增强,鳔充气,显微镜下呈亮黑色,紧贴位于腹腔背部,胃肠蠕动明显,尾鳍鳍膜变宽,尾部色素带消失,全长、体高增加明显(图4.5、图4.6);饥饿组仔鱼身体出现生理性萎缩,头大身细,下颌尖突,脑部凹陷,肠道萎缩,部分仔鱼脊椎弯曲,胃肠内吞入较多气泡(图4.8、图4.9),仔鱼贴壁或沉底,活动能力差,丧失捕食能力。
3 讨论
3.1 仔鱼的混合营养期
鱼类仔鱼从开口摄食到建立完善的外源性营养之前一般要经历混合营养阶段。不同鱼类仔鱼开口时间及混合营养期长短也不同。豹纹鳃棘鲈()仔鱼在2.5日龄开口摄食,3日龄内源性营养耗尽,混合营养期仅0.5 d (杨育凯等, 2017);而大西洋鳟()仔鱼在41日龄开口摄食,71日龄卵黄囊吸收完毕,混合营养期长达30 d (Hansen, 1985);这显然与它们的卵质大小、胚胎发育类型及卵黄囊吸收速度等有关。黑棘鲷初孵仔鱼卵黄囊体积为(0.1997±0.0176) mm3(=10),属于少卵黄型鱼类,仔鱼孵化后卵黄囊吸收迅速,第1天即消耗总体积的70%,至3日龄开口时卵黄囊仅剩最初的1.6%,至6日龄内源性营养耗尽,仅经历了3 d混合营养期,仔鱼必须在短时间内通过捕食建立起外源性营养从而避免进展性饥饿(Progressive starvation)。鱼类的混合营养期除存在种间差异外,还受到发育水温等环境因子的影响。水温低于18℃时,鳀鱼() 2~ 3日龄仔鱼口窝形成,6日龄卵黄消失,混合营养期3~ 4 d;当水温升到23.0℃~24.8℃时,仔鱼2日龄开口,3日龄卵黄消失,混合营养期仅1 d,时间缩短了2~3 d,显然,水温越高,仔鱼的混合营养期越短(万瑞景等, 2004)。为了减小水温和卵黄囊吸收速率等因素对仔鱼混合营养期的种间差异分析的影响,单秀娟等(2008)提出,以仔鱼开口摄食到卵黄囊消耗完毕时间与仔鱼孵化后到开口摄食时间的比率为判别依据,能够比较客观地反映不同鱼类仔鱼混合营养期的相对状况,二者比率>1,说明仔鱼开口摄食较快,不易遭受饥饿胁迫,但多数鱼类该比率<1,仔鱼早期摄食能力低下和遭受饥饿的风险较高。鲷科鱼类大多属于第2种类型:真鲷()仔鱼2日龄开口,4~5日龄卵黄囊消失(鲍宝龙等, 1998);金头鲷()仔鱼3日龄开口,5日龄卵黄囊消失(Yúfera, 1993);平鲷() (周宏团等, 1990)、黄鲷()(夏连军等, 2004)和花尾胡椒鲷() (黄良敏等, 2003)仔鱼都是在开口当天卵黄囊即消失,比率远<1。本研究中,黑棘鲷仔鱼相对混合营养期比率为0.33<1,说明其容易遭受饥饿威胁,仔鱼需要相对较长的时间来提高摄食能力和捕食经验。因此,在实际生产中,应及时投喂饵料,避免仔鱼长期处于饥饿状态而造成机体受损。
3.2 仔鱼初次摄食率和PNR
通常用仔鱼初次摄食率的高低和最高摄食率持续时间的长短来判断其摄食能力。根据初次摄食率的高低变化,可将鱼类分为A和B两种类型:A型仔鱼如金鲫(red variety) (许郑超等, 2015)、花鲈() (王晓龙等, 2017)等,开始时初次摄食率较低,然后迅速上升,再迅速下降;B型仔鱼如半刺厚唇鱼() (秦志清等, 2015)、长薄鳅() (梁银铨等, 2015)等,开始时初次摄食率较低,然后迅速上升,在最高值保持一段时间后再迅速下降,显然 B型仔鱼能够开口摄食的时间更久,摄食能力更强。仔鱼的初次摄食率与口裂大小、游泳能力以及器官发育进程等因素有关,同时,也受到饵料密度以及水生态条件的影响。黑棘鲷属于A型鱼类,口裂较小,游泳能力一般,仔鱼3日龄开口时初次摄食率仅为30%,5日龄初次摄食率达到最高(90%),之后迅速下降,7日龄初次摄食率降至45%,8日龄仅为15%,仔鱼在最高初次摄食率持续时间很短,摄食能力较差。
PNR是评价鱼类饥饿耐力的重要指标之一,到达PNR的时间越长,仔鱼耐受饥饿的能力越强,反之则越弱(黄海等, 2012)。黑棘鲷PNR发生在7日龄,与斑鳜()(王茂元, 2015)、褐菖鲉()(邱成功等, 2014)相似,而齐口裂腹鱼仔鱼的PNR出现在19~20日龄(董艳珍等, 2018)、杂交鲟(Linnaeus×)为25~26日龄(Song, 2004)、哲罗鱼()为39~40日龄(张永泉等, 2009),显然,黑棘鲷PNR发生较早,仔鱼耐受饥饿的能力较弱。在对众多鱼类的研究发现,仔鱼抵达PNR的时间具有随温度升高而缩短的特点,水温越高,仔鱼代谢越快,内源性营养消耗也越快,从而导致外源性营养阶段的提前,最终结果是PNR时间的提前(Bisbal, 1995; 李秀玉等, 2005; 孙中之等, 2009)。为了在比较不同鱼类PNR时具有更加合理的标准,近年来,有学者在研究PNR时采用有效积温(Sum of effective temperature)的概念,利用该参数可以较客观地比较不同鱼类的饥饿忍耐能力,其计算公式可表示为:实验过程平均水温(℃)×PNR时间(d) (Dou, 2002)。多数种类仔鱼PNR有效积温范围为100~250℃·d (陈国柱等, 2007),本研究中,黑棘鲷PNR有效积温为129.5℃·d,处在较下游的位置。黑棘鲷的PNR特点是其在长期进化过程中形成的对外界饵料环境的适应性策略。
3.3 饥饿仔鱼的生理适应机制
鱼类仔鱼在遭受饥饿胁迫时,为了提高存活和摄食的几率,在其生长、发育方面通常会出现一系列的适应性反应。黑棘鲷生长情况与Farris (1959)对仔鱼早期生长划分的3个相期基本相符:仔鱼在孵化后的内源性营养期生长迅速,开口至卵黄囊消失前后长速放缓,6日龄之后对照组仔鱼摄食外源营养进入稳定生长期,饥饿组则出现生长停滞和小幅生理性萎缩,2组全长差异越来越显著(<0.05)。早期仔鱼的个体生长与其发育程度密切相关,在本研究中,饥饿组黑棘鲷仔鱼油球吸收速率明显慢于正常投喂的对照组,原因是饥饿能延缓仔鱼对内源性营养的利用速率,以此来延长饥饿仔鱼的存活时间,让其有更大机会去开口摄食,这是卵黄囊期仔鱼生长的一种自我保护机制,可以在饵料缺少的情况下,提高仔鱼的成活率。随着饥饿加剧,仔鱼为了存活,各组织器官被代谢消耗,导致器官萎缩,身体畸形,待自身储备能量不足时,生长就会受到抑制,鱼体出现生理性萎缩,这是骨骼系统尚未发育的仔鱼为保障活动耗能,提高摄食存活机会的一种生存策略(曲焕韬等, 2017)。
饥饿对仔鱼后期的影响主要表现在形态和行为方面。黑棘鲷饥饿组仔鱼出现明显胸角,并表现出头部畸形、脊椎弯曲、眼球突出、肠道萎缩等一系列明显因饥饿所致的形态学特征。在自然状态下,黑棘鲷仔鱼为了躲避敌害、提高种群成活率,形成了高度集群的习性。本研究中,黑棘鲷集群数量与饥饿时间呈负相关,仔鱼开口后大量集群,饥饿2 d即有1/2离群分散游动,3 d后不再有集群行为,饥饿驱使仔鱼为寻求更多摄食机会而离群,在对豹纹鳃棘鲈(杨育凯等, 2017)、斑鳜(王茂元, 2015)、哲罗鱼(张永泉等, 2009)等鱼类研究中均发现类似现象。鱼类的集群和离群之间存在替代效应,即集群带来的受保护利益与离群带来的获得更多摄食机会之间的替代型转换,饥饿使黑棘鲷集群性减弱符合动物在进化过程中其适合度最大化原则(李彩娟等, 2016)。巡游模式的建立是仔鱼进入外源性营养阶段的重要标志,对照组黑棘鲷仔鱼开口后逐渐开始巡游式运动并建立外源性营养,饥饿组黑棘鲷因缺少食物,仔鱼在卵黄囊消失后表现出烦躁不安并四处狂游,随着饥饿时间的延长,游动速度逐渐减慢,直至静止、死亡。在饥饿的刺激下,仔鱼以增加游动的方式来增加同食物的相遇频率,这体现了仔鱼潜在的生理适应性;但同时也消耗了更多的能量,加速了仔鱼的死亡。因此,在生产实践中,对于耐受饥饿能力较弱的种类,应在仔鱼进入外源性营养时期之前及时投喂饵料,使其积累捕食经验和建立巡游模式,从而提高成活机率(于欢欢等, 2015)。
Bao BL, Su JX, Yin MC. Effect of delayed feeding on feeding ability, survival and growth of red sea bream and olive flounder larvae during early development. Journal of Fisheries of China, 1998, 22(1): 33–38 [鲍宝龙, 苏锦祥, 殷名称. 延迟投饵对真鲷、牙鲆仔鱼早期阶段摄食、存活及生长的影响. 水产学报, 1998, 22(1): 33–38]
Bisbal GA, Bengtson DA. Effects of delayed feeding on survival and growth of summer flounderlarvae. Marine Ecology Progress Series, 1995, 121(1): 301–306
Blaxter JHS, Hempel G. The influence of egg size on herring larvae (L.). Journal du Conseil, 1963, 28(2): 211–240
Chen GZ, Fang ZQ. Effects of starvation on feeding and growth of the larvae of. Chinese Journal of Zoology, 2007, 42(5): 49–61 [陈国柱, 方展强. 饥饿对唐鱼仔鱼摄食和生长的影响. 动物学杂志, 2007, 42(5): 49– 61]
Chen SY, Zhang ZY, Xu J,. Study on larval early development and point of no return of hybrid F2of(♀)×(♂). South China Fisheries Science, 2018, 14(4): 122–128 [陈淑吟, 张志勇, 许津, 等. 黑鲷(♀)×真鲷(♂)杂交子二代仔鱼早期发育及饥饿不可逆点. 南方水产科学, 2018, 14(4): 122– 128]
Dong YZ, Deng SH, Xiao WY,. Effects of delayed feeding on foraging, growth and survival oflarvae.Fisheries Science and Technology Information, 2018, 45(2): 61–64 [董艳珍, 邓思红, 肖文渊, 等. 延迟投饵对齐口裂腹鱼仔鱼摄食, 存活和生长的影响. 水产科技情报, 2018, 45(2): 61–64]
Dou S, Masuda R, Tanaka M,. Feeding resumption, morphological changes and mortality during starvation in Japanese flounder larvae. Journal of Fish Biology, 2002, 60(6): 1363–1380
Duan, GQ, Jiang H, Hu YT,. Experimental starvation onlarvae and goldlarvae and definition of the point of no return. Journal of Aquaculture, 2018, 39(4): 31–35 [段国庆, 江河, 胡玉婷, 等. 鳜和金色鳜仔鱼早期发育与不可逆生长点确定. 水产养殖, 2018, 39(4): 31–35]
Farris DA. A change in the early growth rates of four larval marine fishes. Limnology and Oceanography, 1959, 4(1): 29–36
Gao XQ, Hong L, Liu ZF,. The definition of point of no return of larvae and feeding characteristics oflarvae and juveniles. Journal of Fisheries of China, 2015, 39(3): 392–400 [高小强, 洪磊, 刘志峰, 等. 美洲西鲱仔鱼不可逆点及仔、稚鱼摄食特性研究. 水产学报, 2015, 39(3): 392–400]
Hansen TJ, Møller D. Yolk absorption, yolk sac constrictions, mortality, and growth during first feeding of Atlantic salmon () incubated on astro-turf. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(6): 1073–1078
Huang H, Yang N, Zhang X. Experimental starvation onlarvae and determination of point of no return. South China Fisheries Science, 2012, 8(2): 43–50 [黄海, 杨宁, 张希. 马拉瓜丽体鱼仔鱼饥饿实验及不可逆点的确定. 南方水产科学, 2012, 8(2): 43–50]
Huang LM, Xie YJ, Deng SP,. Effects of delayed initial feeding on foraging, growth and survival of larval yellow spotted grunt(). Journal of Jimei University (Natural Science), 2003, 8(2): 130–133 [黄良敏, 谢仰杰, 邓书品, 等. 延迟投饵对花尾胡椒鲷仔鱼摄食、生长和存活的影响. 集美大学学报(自然科学版), 2003, 8(2): 130–133]
Li CJ, Xu ZC, Zhang ZZ,. Starvation and point of no return oflarvae. Journal of Yangzhou University (Agricultural and Life Science), 2016, 37(3): 65–70 [李彩娟, 许郑超, 张振早, 等. 赤眼鳟仔鱼饥饿实验和不可逆点研究. 扬州大学学报(农业与生命科学版), 2016, 37(3): 65–70]
Li XY, Lin X, Liao ZH,. Effect of temperature on feeding oflarvae. Ecologic Science, 2005, 24(3): 243–245 [李秀玉, 林小涛, 廖志洪, 等. 温度对黄颡鱼仔鱼摄食强度及饥饿耐受力的影响. 生态科学, 2005, 24(3): 243–245]
Liang YQ, Chen J, Fang YH,. Effects of delayed initial feeding onlarvae. Freshwater Fisheries, 2015, 45(2): 107–110 [梁银铨, 陈静, 方艳红, 等. 延迟初次摄食对长薄鳅仔鱼的影响. 淡水渔业, 2015, 45(2): 107–110]
Qin ZQ, Liu YJ, Fan HP,. Effects of delayed initial feeding on foraging, growth and survival oflarvae. Journal of Fujian Fisheries, 2015, 37(5): 392–398 [秦志清, 刘亚君, 樊海平, 等. 延迟投饵对半刺厚唇鱼仔鱼摄食、生长与存活的影响. 福建水产, 2015, 37(5): 392–398]
Qiu CG, Xu SL, Lin SZ,. Effects of delayed feeding on survival and growth oflarva. Journal of Ningbo University (NSEE), 2014, 27(3): 7–11 [邱成功, 徐善良, 林少珍, 等. 延迟投喂对褐菖鲉仔鱼存活和生长的影响. 宁波大学学报(理工版), 2014, 27(3): 7–11]
Qu HT, Liu Y, Hu MH,. Effects of starvation on feeding, growth and development oflarvae. Chinese Journal of Fisheries, 2017, 30(6): 24–29 [曲焕韬, 刘勇, 胡美洪, 等. 饥饿对圆口铜鱼仔鱼早期发育的影响. 水产学杂志, 2017, 30(6): 24–29]
Shan XJ, Dou SZ. Effects of delayed first feeding on growth, survival and biochemical composition of croakerlarvae. Oceanlogia et Limnologia Sinica, 2008, 39(1): 14–23 [单秀娟, 窦硕增. 饥饿胁迫条件下黑鮸()仔鱼的生长与存活过程研究. 海洋与湖沼, 2008, 39(1): 14–23]
Song B, Chen LQ, Gao LJ,. The effects of starvation on feeding, growth and biochemical composition of hybrid sturgeon larvae. Acta Hydrobiologica Sinica, 2004, 28(3): 333–336
Sun ZZ, Liu XZ, Xu YJ,.Effects of starvation on the growth and development of larval and juvenile rock bream. Progress in Fishery Sciences, 2009, 30(4): 8–13 [孙中之, 柳学周, 徐永江, 等. 饥饿对条石鲷仔稚鱼生长发育的影响. 渔业科学进展, 2009, 30(4): 8–13]
Wan RJ, Li XS, Zhuang ZM,. Experimental starvation onlarvae and definition of the point of no return. Journal of Fisheries of China, 2004, 28(1): 79–83 [万瑞景, 李显森, 庄志猛, 等. 鳀鱼仔鱼饥饿实验及不可逆点的确定. 水产学报, 2004, 28(1): 79–83]
Wang MY. Starvation test onlarvae and acertaining the point of no return. Journal of Guangdong Ocean University, 2015, 35(4): 99–103 [王茂元. 斑鳜仔鱼饥饿实验及不可逆点的确定. 广东海洋大学学报, 2015, 35(4): 99–103]
Wang XL, Wen HS, Zhang MZ,. Determination of irreversible starvation point and feeding rhythm oflarvae. Periodical of Ocean University of China, 2017, 47(5): 57–64 [王晓龙, 温海深, 张美昭, 等. 花鲈初孵仔鱼饥饿不可逆点的确定及摄食节律研究. 中国海洋大学学报, 2017, 47(5): 57–64]
Wu RX, Liu J, Fan JR,. A review on the nomenclature and taxonomic status of the black porgy,(Perciformes: Sparidae). Marine Sciences, 2011, 35(5): 117–119 [吴仁协, 刘静, 樊冀蓉, 等. 黑棘鲷的命名和分类地位探究. 海洋科学, 2011, 35(5): 117–119]
Xia LJ, Shi ZH, Lu JX. Experimental starvation onlarvae and definition of the point of no return. Marine Fisheries, 2004, 26(4): 286–290 [夏连军, 施兆鸿, 陆建学. 黄鲷仔鱼饥饿实验及不可逆点的确定. 海洋渔业, 2004, 26(4): 286–290]
Xu XJ, Ou YJ, Li JE. Effects of delayed feeding on foraging, survival and growth of ovate pompano () larvae during early developmental stage. South China Fisheries Science, 2010, 6(1): 37–41 [许晓娟, 区又君, 李加儿. 延迟投饵对卵形鲳鲹早期仔鱼阶段摄食、成活及生长的影响. 南方水产科学, 2010, 6(1): 37–41]
Xu ZC, Wang GC, Liu Q,. Experimental starvation onred variety larvae and determination of the point of no return. Journal of Aquaculture, 2015, 36(4): 14–19 [许郑超, 王国成, 刘青, 等. 金鲫仔鱼的饥饿实验和不可逆点研究. 水产养殖, 2015, 36(4): 14–19]
Yang YY, Yu W, Lin HZ,. Experimental starvation oflarvae and study of point of no return. South China Fisheries Science, 2017, 13(6): 90–96 [杨育凯, 虞为, 林黑着, 等. 豹纹鳃棘鲈仔鱼饥饿实验和不可逆点研究. 南方水产科学, 2017, 13(6): 90–96]
Yin MC. Advances and studies on early life history of fish. Journal of Fisheries of China, 1991, 15(4): 348–358 [殷名称. 鱼类早期生活史研究与其进展. 水产学报, 1991, 15(4): 348–358]
Yu HH, Chen C, Zhang TT,. Feeding and growth oflarvae in the process of nourishing transformation. Progress in Fishery Sciences, 2015, 36(6): 37–42 [于欢欢, 陈超, 张廷廷, 等. 饥饿对云纹石斑鱼 ()卵黄囊期仔鱼摄食和生长的影响. 渔业科学进展, 2015, 36(6): 37–42]
Yúfera M, Pascual E, Polo A,. Effect of starvation on the feeding ability of gilthead seabream (L.) larvae at first feeding. Journal of Experimental Marine Biology and Ecology, 1993, 169(2): 259–272
Zhang YQ, Yin JS, Du J,. Experimental starvation onand definition of the point of no return. Acta Hydrobiologica Sinica, 2009, 33(5): 945–950 [张永泉, 尹家胜, 杜佳, 等. 哲罗鱼仔鱼饥饿实验及不可逆生长点的确定. 水生生物学报, 2009, 33(5): 945–950]
Zhou HT, Li JE, Ma RH,. Observation on embryonic development and morphology of larva, juvenile and young flat bream,. Fisheries Science and Technology Information, 1990(1): 2–5 [周宏团, 李加儿, 马荣和, 等. 平鲷胚胎发育及仔、稚、幼鱼的形态观察. 水产科技情报, 1990(1): 2–5]
Experimental Starvation ofLarvae, and Definition of the Point of No Return
YANG Yukai1, LIN Heizhao1, WANG Zhiyong2①, XIE Yangjie2, GUO Yihui1, HUANG Xiaolin1, LI Tao1
(1. Key Laboratory of South China Sea Fishery Research Exploitation & Utilization, Ministry of Agriculture and Rural Affairs; South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300; 2. Key Laboratory of Healthy Mariculture for the East China Sea, Ministry of Agriculture and Rural Affairs; Fisheries College, Jimei University, Xiamen 361021)
The potential effects of starvation on the growth, feeding, and survival oflarvae were investigated at a water temperature of 18℃~19℃ by using ecological experimental methods. Along with the detection of the initial feeding rate and the point of no return (PNR), the optimum initial feeding time of the larvae were also ascertained. The results showed that larvae started feeding on the third day post hatching (dph), and the yolk sac and the oil globule disappeared at 4 DPH and 6 dph, respectively, which suggested that the duration of the mixotrophic stage was about 3 days and larvae were vulnerable to starvation. When the fish opened their mouths, the initial feeding rate was only 30%, which then rapidly increased to the highest level of 90% at 5 dph, after which it gradually declined to 45% at 7 dph, which denoted the PNR. Significant variation was found between the starved and control groups after 6 dph (0.05). Reduced mobility, distorted or deformed bodies, and increased mortality were observed in the starved larvae after the PNR. All the starved larvae died before 10 DPH. In brief, our study suggested thatlarvae were vulnerable to starvation and the optimum initial feeding time of the larvae was within 4 days after their mouths opened.
; Larvae; Starvation; Point of no return (PNR)
S961
A
2095-9869(2020)02-0087-08
王志勇,教授,E-mail: zywang@jmu.edu.cn
2019-01-04,
2019-01-27
* 农业农村部东海海水健康养殖重点实验室开放课题基金(2013ESHML09)、中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金(2018ZD01)和深圳市科技研发资金项目(JCYJ20170817103856495)共同资助[This work was supported by Open Fund of Key Laboratory of Healthy Mariculture for the East China Sea (2013ESHML09), Central Public-Interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS (2018ZD01), and Shenzhen Science and Technology R & D Funding Project (JCYJ20170817103856495)]. 杨育凯,E-mail: yangyukai1986@163.com
10.19663/j.issn2095-9869.20190104001
http://www.yykxjz.cn/
杨育凯, 林黑着, 王志勇, 谢仰杰, 郭奕惠, 黄小林, 李涛. 黑棘鲷仔鱼饥饿实验及不可逆点的确定. 渔业科学进展, 2020, 41(2): 87–94
Yang YK, Lin HZ, Wang ZY, Xie YJ, Guo YH, Huang XL, Li T. Experimental starvation oflarvae, and definition of the point of no return. Progress in Fishery Sciences, 2020, 41(2): 87–94
WANG Zhiyong, E-mail: zywang@jmu.edu.cn
(编辑 马璀艳)
