陆地棉COR基因在低温胁迫下的表达分析及功能鉴定
2020-03-18李建平足木热木吐尔逊郝晓燕常晓春高升旗胡文冉黄全生
李建平,足木热木·吐尔逊,郝晓燕,常晓春,高升旗,胡文冉,陈 果,黄全生
(新疆农业科学院核技术生物技术研究所/新疆农作物生物技术重点实验室,乌鲁木齐 830091)
0 引 言
【研究意义】 在植物的整个生长周期中,干旱、低温和盐碱等非生物因素限制其生长发育和生物产量。为了应对和适应这些不利的环境因素,植物从生化和生理层面不断的进行自我进化[1]。当前,发掘非生物胁迫诱导基因及其信号通路相关组分的工作取得了显著进展,发掘出许多与非生物胁迫相关的转录因子和蛋白,如与干旱胁迫、高盐胁迫等相关的钙依赖蛋白激酶(CDPK)家族[2],ICE、CBF /DREB1及冷调控基因COR(cold regulated gene)家族与低温胁迫有关[3,4],促分裂素原活化蛋白激酶(Mitogen-activated protein kinases, MAP激酶,MAPK)则在生物胁迫和非生物胁迫通路中均发挥功能[5,6]。不同的非生物胁迫因子对植物的生长发育所造成的影响是普遍而又特殊的[7]。近几年,围绕非生物胁迫反应的调控生理机制,胁迫途径相关功能基因、转录因子等的发掘及利用基因工程技术提高棉花的抗胁迫能力的研究工作逐步开始大量开展[8]。尽管棉花抗逆基因克隆和抗逆转基因棉花研究取得了一定进展,尤其一些冷胁迫相关基因被克隆,但其提高棉花低温胁迫的作用机制还知之甚少。【前人研究进展】棉花一般适合在温暖的地区生长[9],低于15℃的温度会严重影响棉花的生长发育,使棉株容易感染真菌而诱发多种病症,大田缺苗断垄,导致减产[10]。棉花在整个生育期均容易遇到低温胁迫,主要是在萌发期和苗期。棉花生长季节早期的低温会对棉花幼苗造成严重伤害,在温度恢复正常后棉株也可能无法完全恢复。Zhao 等[11]研究表明,短期低温胁迫可能造成棉花叶片一定程度的生理障碍,当这些经低温胁迫(白天16℃/ 夜间12℃或白天12℃/ 夜间8℃处理3 d) 后受伤的叶片进一步接种链格孢菌时,棉花叶片出现叶斑病的明显症状并呈发展趋势,最终出现明显的衰老症状。传统育种方式对于新品种的培育周期过长,目前尚未培育出耐冷性能突出的新品种。近年来,新一代测序(Next generation sequencing,NGS)技术已应用于全基因组表达谱研究[12-13]。Fernandez 等[14]利用转录组对参与向日葵响应低温胁迫的候选基因进行了分析,有80个基因响应早期的冷胁迫,这些基因分别拥有不同的功能-转录、翻译、蛋白质降解、蛋白质折叠、ROS产生、ROS清除等。Park等[15]在转录组水平研究表明,棉花受水分胁迫后,519个转录本表现出差异,其中有147个转录本有功能注释,这些转录本中有70%参与以下4类功能中,分别为胁迫/ 防御型、代谢型、基因调控型及未明确分类型,其中热激蛋白相关转录本和活性氧相关转录本最丰富。表明多倍体的棉花受水分胁迫后的转录组响应涉及复杂的机制。国内外利用分子生物学方法已经克隆到一些棉花逆境胁迫响应基因。Ma等[19]克隆了GhCBF基因,发现受到低温胁迫诱导。2013年Xu等[20]发现来源于棉花的GhCAX3 在酵母和烟草中表达能显著提高其抗冷性。Kargiotidou等[21]发现棉花FAD2-3和FAD2-4受到低温诱导。【本研究切入点】尽管棉花抗逆基因克隆和抗逆转基因棉花研究取得了一定进展,尤其一些冷胁迫相关基因被克隆,但其提高棉花低温胁迫的功能还知之甚少,尤其是陆地棉COR基因在棉花抗低温胁迫中的功能研究目前尚鲜见报道。研究陆地棉COR基因在低温胁迫下的表达分析及功能鉴定。【拟解决的关键问题】基于前人研究的结果及陆地棉基因组数据库,发掘与棉花耐低温相关的COR候选基因,进行克隆及功能验证,研究其作用机理,通过遗传改良的方式培育耐冷棉花新品种。
1 材料与方法
1.1 材 料
陆地棉标准系TM-1。
1.1.1 菌株与载体
大肠杆菌TransT1购自全式金生物,农杆菌菌株GV3101为实验室保存。克隆载体pEASY Blunt vector购自全式金生物, VIGS病毒载体由南京农业大学惠赠。
1.1.2 试剂
所用的 PRIMESTAR DNA聚合酶购自Takara公司;TaqDNA聚合酶购自康为公司;M-MLV购自Invitrogen公司,各类限制性内切酶及连接酶购自NEB公司;Trizol购自Invitrogene公司。Real-time PCR试剂盒购自ABI公司。
1.1.3 引物设计
陆地棉COR基因序列号及mRNA序列基于陆地棉基因组数据库(https://www.cottongen.org/)及国际生物信息库(https://www.ncbi.nlm.nih.gov/);Real-time PCR引物及VIGS引物设计基于Primer Premier 5.0完成。表1
1.2 方 法
1.2.1 棉苗种植及低温处理
经硫酸脱绒的棉籽置于用水浸湿的发芽纸上并用另一浸湿的发芽纸覆盖密封,置于26~28℃的温箱中放置4~6 d带胚芽从棉籽中露出。在8.0×8.0(cm)的方形培养钵(商品化)中装满花卉营养土,并用自配Hoagland营养液浸透,将出芽状况近似的棉种埋于营养土下约3~4 cm处。在23~25℃ 16h Light/8h Dark光周期下培养7~10 d至棉苗子叶展开但尚未长出真叶。按照一个处理每个品种种植30株,共计3个处理,在26℃ 16 h Light/8h Dark光周期下培育3周至发育出2~3片真叶,然后转至10℃ 16 h Light/8h Dark处理0、3、6、12、24和48 h。分别在每个时间点取样,提取总RNA备用。将3周苗龄的棉苗移至10℃进行低温胁迫处理。以低温处理前的样品为对照,分别在处理3、6、12、24及48 h时采样,经过总RNA提取、cDNA合成及实时荧光定量PCR等步骤,对候选基因受低温诱导时的基因表达情况进行分析。
1.2.2 cDNA第一链合成依据Invitrogen公司M-MLV使用说明操作
Real-time PCR检测依据ABI公司Real-time PCR试剂盒使用说明操作。
1.2.3 VIGS载体构建和转化
根据Gh_D12G0215目的基因序列,在其保守区域选取一段300~600 bp的序列设计引物,并在正向引物5’端引入EcoRI酶切位点,反向引物5’端引入KpnI酶切位点,引物序列:Gh_D12G0215-VF:GGAATTCCTTCTCATCAGTTTTCTATCCT;Gh_D12G0215-VR:GGGGTACCGTAATGTGTGTCAGACTTCGG(下划线部分为引入的酶切位点)。以棉花总RNA为模板,通过RT-PCR克隆目的片段,连接至pEASY Blunt vector并测序。将测序正确的克隆利用EcoRI与KpnI切酶从pEASY Blunt vector双酶切并回收,连接至同样经EcoRI与KpnI酶切后的VIGS载体上,转至GV3101农杆菌中保存备用。
以棉花cDNA为模版,PCR扩增得到候选COR基因成员Gh_D12G0215的VIGS目的片段,将该片段回收后连接至pEASY blunt克隆载体上送测序公司测序。选取测序结果与原始序列完全一致的克隆提取质粒,同时提取VIGS病毒载体质粒。用限制性内切酶EcoRⅠ与KpnⅠ双酶切克隆载体和VIGS载体并回收。以T4DNA连接酶连接目的片段至VIGS载体,挑取阳性克隆并酶切鉴定。
1.2.4 目的基因沉默
VIGS操作方法详见Gao 等[19]。
作为对照,将TRV:GhCLA1与TRV:RNAi共同侵染棉花子叶后,将棉苗在23℃避光环境下培养24 h,再转至同样温度下12 h光照,12 h黑暗光周期下继续生长7~10 d至真叶长出,观察新长出的真叶是否出现白化表型,进行的基因沉默操作结果是有效的。将基因沉默后的棉苗与对照共同在10℃下进行低温胁迫处理,观察棉苗在低温条件下的生长表现,并且将棉苗在低温处理48 h后移至26℃恢复生长72 h统计存活率。
1.2.5 基因功能鉴定
将VIGS处理并已出现沉默效应的棉苗移至10℃ 16 h Light/8 h Dark处理48 h,观察表型并统计。
表1COR基因家族引物信息
Table 1 The information ofCORfamily member primers
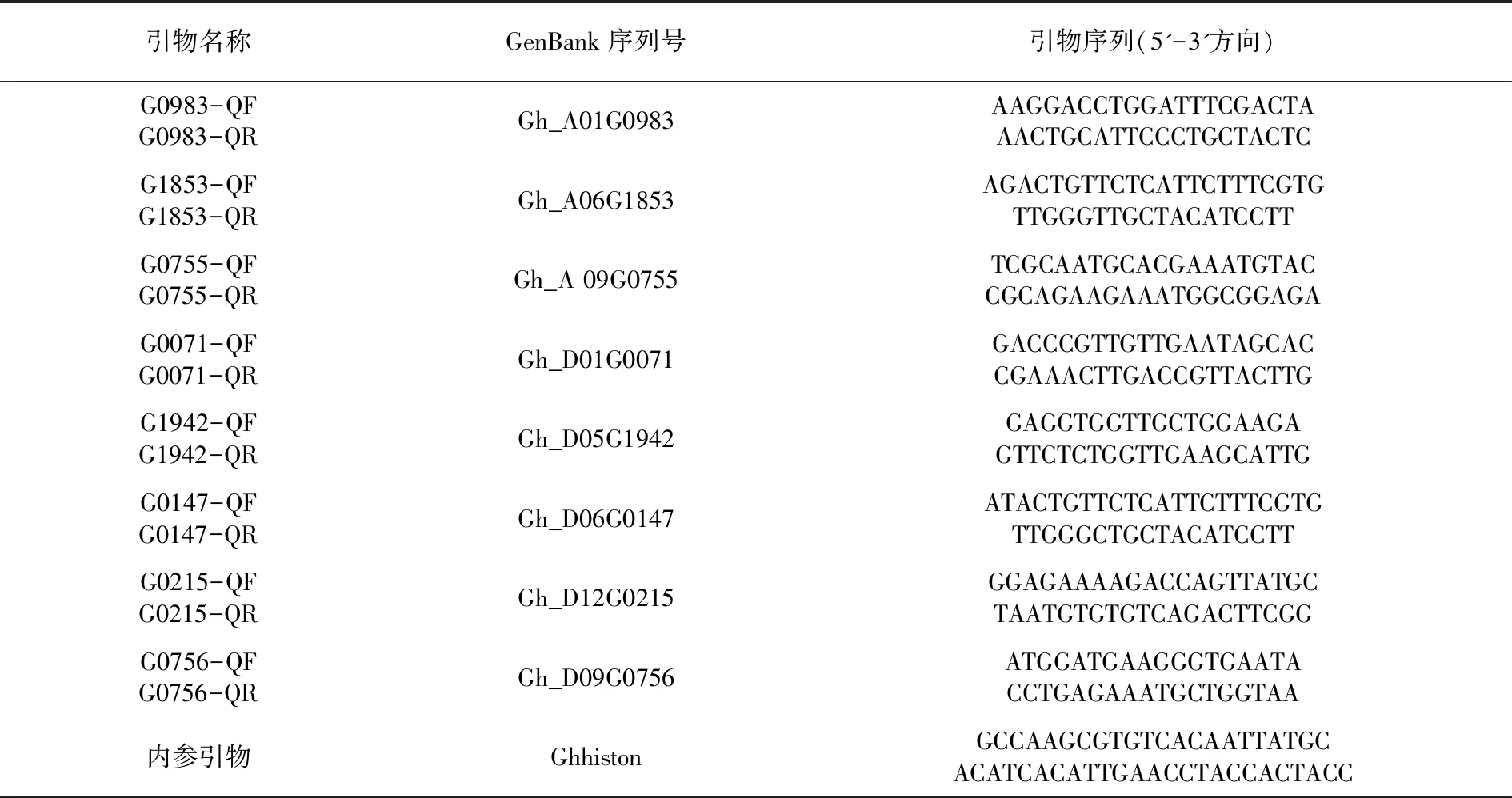
引物名称GenBank序列号引物序列(5'-3'方向)G0983-QFG0983-QRGh_A01G0983AAGGACCTGGATTTCGACTA AACTGCATTCCCTGCTACTCG1853-QFG1853-QRGh_A06G1853AGACTGTTCTCATTCTTTCGTGTTGGGTTGCTACATCCTTG0755-QFG0755-QRGh_A 09G0755TCGCAATGCACGAAATGTACCGCAGAAGAAATGGCGGAGAG0071-QFG0071-QRGh_D01G0071GACCCGTTGTTGAATAGCACCGAAACTTGACCGTTACTTGG1942-QFG1942-QRGh_D05G1942GAGGTGGTTGCTGGAAGAGTTCTCTGGTTGAAGCATTGG0147-QFG0147-QRGh_D06G0147ATACTGTTCTCATTCTTTCGTGTTGGGCTGCTACATCCTTG0215-QFG0215-QRGh_D12G0215GGAGAAAAGACCAGTTATGCTAATGTGTGTCAGACTTCGGG0756-QFG0756-QRGh_D09G0756ATGGATGAAGGGTGAATACCTGAGAAATGCTGGTAA内参引物GhhistonGCCAAGCGTGTCACAATTATGCACATCACATTGAACCTACCACTACC
2 结果与分析
2.1 陆地棉COR基因表达受低温胁迫诱导
研究表明,部分COR基因家族成员响应低温诱导,基因表达发生不同程度的变化。基因序列号为Gh_D06G0147的COR基因受低温胁迫基因表达受到显著抑制,该基因在低温胁迫中可能起负调控的作用;Gh_A09G0755在胁迫初期至6 h,基因表达上调了2倍,但是随着胁迫时间的延伸,表达水平出现下降的趋势,但是,在胁迫48 h后仍然达到对照的1.25倍,该基因可能参与了低温胁迫调控;Gh_D12G0215在低温胁迫作用下,随着低温处理的时间延长,基因表达水平逐步升高,在处理48 h后,表达水平达到对照的9.1倍,该基因受低温胁迫诱导基因表达非常明显,可能在棉花低温逆境条件下发挥一定的作用。其余COR基因家族成员的表达收到低温胁迫的影响和对照相比无显著的变化,但是并不表明这些基因的功能较弱,可能这些基因在其他逆境中如高温、盐胁迫中发挥作用。图1
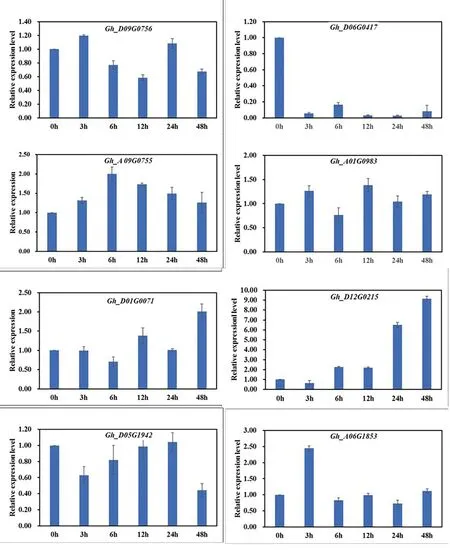
图1 陆地棉COR基因家族成员低温诱导下的表达
Fig.1 Relative expression analysis of differentCORgene family members induced by low temperature
2.2 COR基因的VIGS载体构建
包含COR基因部分序列的VIGS载体构建成功。将构建好的载体转至农杆菌菌株GV3101中保存备转化棉苗用。图2
2.3 病毒诱导的COR基因沉默导致棉花对低温胁迫更为敏感
将TRV:Gh_D12G0215与TRV:RNAi共同侵染棉花子叶,使棉花中的内源COR基因Gh_D12G0215发生沉默。通过Real-time PCR检测发现GhCLA1及Gh_D12G0215的表达水平降低,该基因在棉花内已被沉默。
研究表明,在相同的处理条件下,基因沉默后的棉苗存活率为31.09%,对照为53.27%,与对照相比,发生基因沉默的棉苗对低温胁迫更为敏感,Gh_D12G0215基因可能参与调控棉花在低温胁迫下的信号传递途径。图3

注:M为Trans2K DNA Marker,1为TRV:RNA2 空载体,2为TRV:Gh_D12G0215经EcoRI与KpnI双酶切后结果。载体片段为3 180 bp,目的片段大小为350 bp
Note: M, Trans2K DNA Marker; 1, TRV:RNA2 empty vector; 2, TRV:Gh_D12G0215 digested byEcoRIandKpnI.The length of vector fragment is 3,180 bp, and the length of target fragment is 350bp
图2 TRV:Gh_D12G0215载体酶切鉴定
Fig. 2 The Double digested by EcoRI and KpnI result of TRV:Gh_D12G0215 construction vector
3 讨 论
陆地棉是异源四倍体,基因组预测分析显示,大约含7万个基因[20],而雷蒙德氏棉基因组中约含3.7万基因[21],需要建立高通量、准确、易操作体系快速揭示基因功能。2008年,Gao等[22]通过VIGS方法抑制棉花抗黄萎病相关基因GhNDR1和GhMKK2表达, 阐明了其在棉花抗病研究中利用潜力。近几年,该技术在基因的功能研究方面被广泛使用并得到国际认可,故而VIGS技术具有简单、快速、高通量、不受遗传背景影响等优点,利用VIGS体系可高通量研究棉花基因功能,特别是快速查明表型明显,作用大的重要质量性状基因,为目标性状基因的快速发掘与育种应用提供重要基因资源与依据。研究中,同样利用实验室建立的VIGS技术体系,验证了陆地棉COR基因在棉花受低温胁迫时发挥一定的功能。研究中,除Gh_D12G0215的表达受低温胁迫上调外,其它基因的表达或下调,或表达量出现短暂提高进而有所降低,但是棉花中的其余COR家族成员是否在低温胁迫或其它逆境胁迫中发挥一定的功能仍有待证明。

图3 陆地棉Gh_D12G0215基因沉默植株在低温胁迫下的表型及存活率
Fig. 3 Phenotype of silencedGh_D12G0215 seedlings after low temperature treatment
4 结 论
陆地棉COR基因家族受低温胁迫基因表达各不相同,序列号为Gh_D06G0147的COR基因受低温胁迫基因表达受到显著抑制,基因表达水平低于对照,暗示该基因在低温胁迫中可能起负调控的作用,但是实际功能有待进一步验证;Gh_A09G0755在胁迫初期至6 h,基因表达上调了2倍,但是随着胁迫时间的延伸,表达水平出现下降的趋势,在胁迫48 h后仍然达到对照的1.25倍,该基因可能参与了低温胁迫调控;Gh_D12G0215在低温胁迫作用下,随着低温处理的时间延长,基因表达水平逐步升高,在处理48 h后,表达水平达到对照的9.1倍,基因序列号为Gh_D12G0215的COR基因在棉花中沉默后,棉花对低温胁迫表现的更为敏感,暗示着该基因可能参与了陆地棉在低温胁迫下的调控。序列号为Gh_D12G0215的COR基因在棉花中沉默后,在低温条件下的存活率仅为31.09%,显著低于对照的存活率(对照存活率为53.27%)。通过表型观察和统计,确定Gh_D12G0215参与了低温胁迫下的调控。
