芦芽山针叶林分布上下限土壤温度及含水量的季节差异
2020-03-13王明昌董满宇丁新原杨显基崔明皓康慕谊
薛 峰,江 源,*,王明昌,董满宇,丁新原,杨显基,崔明皓,康慕谊
1 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 2 北京师范大学地理科学学部, 北京 100875
土壤温度和水分含量作为土壤重要物理特性,在根系生长[1]、种子萌发[2]、土壤有机质分解[3]和养分元素迁移[4]等一系列生物化学过程中发挥着重要作用。同时,土壤温度和水分含量也深刻影响着土壤动物与微生物的数量和多样性以及土壤酶的活性[5- 7],进一步影响到地上生态系统中植物的生长及地理分布[8- 9]。
区域尺度上,土壤水热状况和动态变化主要取决于纬度、大陆度及土地利用类型[10- 11];小尺度下,土壤温度和含水量直接受地形、辐射、凋落物厚度和土壤质地等因子的影响[12- 13]。比如,祁连山和甘南草甸的研究表明,阳坡土壤温度是阴坡的数倍之高,同时土壤含水量却不及阴坡的一半[14- 15]。土壤温度和含水量之间还存在交互影响。水的热容量远大于土壤固态物,土壤中水分的相变和含量动态影响土壤热容量和导热率,而土壤温度的变化也会造成水分的蒸发和凝结[16- 17],这种耦合关系在高山高寒地区土壤周期性冻融过程中尤其重要。因此,我国土壤温湿度动态的相关研究主要集中在青藏高原高寒草甸和高山林线。李卫明等通过分析藏北高原冻融过程中土壤温度日尺度和季节尺度变化特征,发现观测期日尺度上土壤温度变化缓和,冬春季土壤温度变幅剧烈,反映了冻融过程土壤含水量变化对土壤温度的显著调节作用[18];俞洁辉等通过对唐古拉山高山草甸海拔分布上限土壤温湿度连续3a的季节变化分析,揭示了限制草甸向更高海拔分布的原因[19]。华北地区生态系统深受人类活动影响,相关研究大多关注平原地区农田土壤温湿度变化对农作物生长的影响[20- 21],并且由于高山区气象数据匮乏,有关土壤温湿度研究较少见,多年的系统监测更是几无涉及。
高山林线处于高寒胁迫的临界状态而对气候变化十分敏感,被誉为气候变化的“监视器”[22]。为了揭示高山林线形成的气候机制,最热月均温、生长季平均温度和生长季长度等热量指标和生长指标被用来解释林线的位置[9, 23]。Körner 和 Paulsen 对全球范围内林线林下生态气候的监测研究,发现林线生长季内土壤(10cm)平均温度为6.7℃,而且全球尺度下0.8℃的标准差也说明热量不足对林线树木生长的限制[24]。因此,尽管大量研究表明干旱区、半干旱区和岛屿的高山林线受到季节性干旱的胁迫[25- 26],然而在全球尺度上,土壤水分有效性并不被认为是调节林线树木生长的主要因子[27]。我国海拔在林线以上高度的高山众多,对林线-气候关系研究由来已久[28- 29]。然而,有关林线土壤温湿度特征的研究稍显不足,并且已有的研究结果区域差异性较大。比如,祁连山青海云杉林线生长季土壤均温为4.9℃小于全球均值,而色季拉山方枝柏林线生长季土壤均温(8.0℃)大于全球均值[30- 31]。崔海亭等曾对华北地区高山林线的气候特点和树木生长做过详尽论述,但是由于整个华北地区高山气象站缺乏,仅有五台山气象站在海拔2000m以上,其研究所需的气象数据都是由低海拔气象站推算所得[22]。因此,该研究结果仍待华北地区更多的高山林线土壤温湿度的监测研究等进一步佐证和充实。
位于华北干旱区和半干旱区交界的芦芽山,地处东亚季风区边缘,山体海拔超过林线垂直分布高度,山上成带分布着大面积具水源涵养、气候调节和多样性保育重要作用的寒温性针叶林-青扦(Piceawilsonii)林和白扦(P.meyeri)林[32]。本研究通过对芦芽山南北坡森林分布上限土壤温度和水分连续5a的监测,探讨以下两个问题:(1)芦芽山南北坡林线土壤在周期性冻融过程中温度和含水量的季节变化特征;(2)芦芽山北坡针叶林分布上下限土壤温度和含水量的季节差异性。
1 材料与方法
1.1 研究区概况
研究区位于山西省管涔山脉主峰芦芽山(38°40′—38°50′N, 111°50′—112°00′ E),主峰最高点荷叶坪海拔2772m(图1)。该区地处东亚季风区边缘的暖温带半干旱气候区,大陆性特点显著,夏季凉爽多雨,冬季寒冷干燥。五寨气象站(38°55′N,111°51′E)过去60年(1957—2016)间的气候资料显示,芦芽山地区年均温5.3℃,年降水459.8mm;其中,过去60年该地气温呈显著上升趋势,平均达+0.24 ℃/10a,降水变化则不明显。除2012年外,2013至2016年年均温都超过多年均值,并且2013年降水接近多年最大值(图2)。芦芽山植被分布具有垂直地带性,海拔1300—1800m为暖温带落叶阔叶林和灌草丛带,主要建群种为白桦(Betulaplatyphylla);海拔1950—2600m保存有大面积的寒温性针叶林,其建群种为白扦和华北落叶松(Larixprincipis-rupprechtii),并在海拔2650m左右形成林线;海拔2650m以上开始出现高山草甸[33]。土壤主要发育在残积和/或坡积母质上,对应于植被覆盖,土壤类型随海拔增加依次为山地褐土(栗褐土)、山地淋溶褐土、山地棕壤和亚高山草甸土[34]。
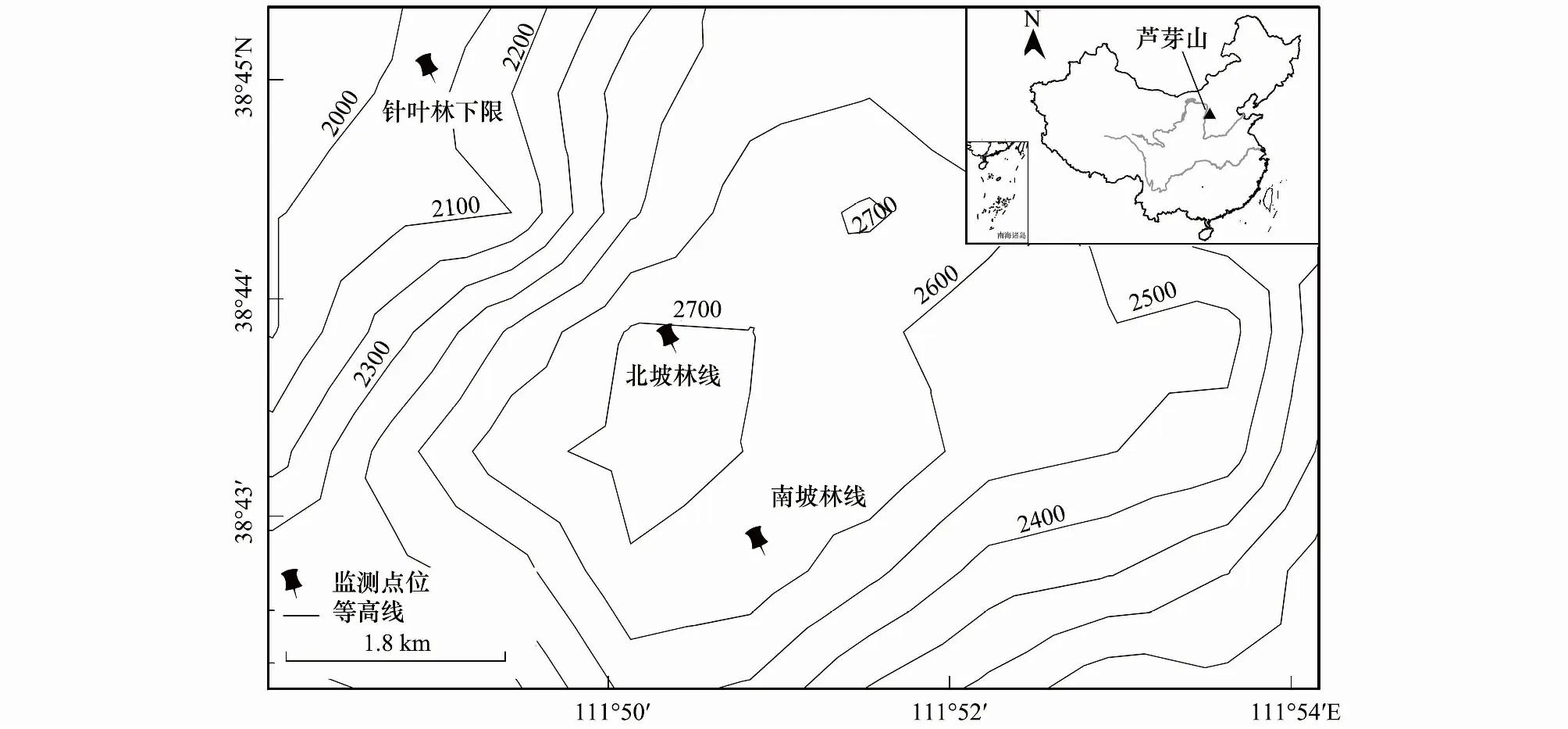
图1 研究区及监测点地理位置Fig.1 Location of the observation sites at Luya Mountain
图2 五寨气象站1957—2016年气温和降水变化状况Fig.2 Changes of air temperature and precipitation from 1957 to 2016 recorded by Wuzhai meteorological station
1.2 样地选择和仪器布设
为监测芦芽山林线土壤温度和含水量的变化状况,2011年8月初,分别在山体的南、北两坡面林线交错带选取理想样地,布设土壤温度和含水量监测仪器。南坡监测点海拔2665m,坡度20°,林冠层盖度约为50%,优势树种为白扦和华北落叶松,树木均高6m;北坡监测点海拔2720m,坡度25°,林冠层盖度为60%,优势树种同为白扦和华北落叶松,树木均高5m。土壤含水量采用土壤水分仪(ML2x, Delta-T, Cambridge, UK)测定,土壤温度采用土壤温度记录仪(ST-05, Delta-T, Cambridge, UK)测定, 利用数据采集器(GP1, Delta-T, Cambridge, UK)进行时间分辨步长为1 record/h(即每1小时记录一次)的不间断数据采集。参照Körner全球林线处温度监测方法[23],仪器探头埋设在树冠投影下土壤深度10cm处,并保证监测点全天不受太阳光照射。仪器机盒做防水包裹处理后,就地掩埋以防丢失,而后逐年定期采集、下载数据并更换电池。本文基于2012/01/01—2016/12/31共5个完整年份的观测数据进行分析。其中2015/12/18 4:00—2016/04/05 10:00因电池故障而导致数据存在部分缺失。
为了与高海拔林线带的土壤温湿数据作对比和交叉印证,我们同时在芦芽山北坡针叶森林分布的下边界(海拔2040 m)埋设仪器,监测土壤温湿度变化。监测点优势树种和土壤类型与南北坡林线处大体一致,只是坡度(10°)较缓,郁闭度(90%)和树高(9 m)明显更大。
1.3 数据处理和分析
首先,根据24小时实测数据计算土壤温度日均值和日较差
DRST=Hmax-Hmin
式中,Td为土壤日均温,DRST为土壤温度日较差,Hi为一天内中i时刻的实测土壤温度,Hmax为一天中土壤最高温,Hmin为最低温。相应的,我们根据土壤日均值计算得到土壤温度的月均值和年均值,
式中,TY和TM分别为土壤年均温和月均温,Y为当年的天数,M为当月的天数。为了进一步反映土壤温度日较差的季节变化,根据日较差数据进一步求得其月均值
式中,MRST为土壤温度日较差的月均值,DRSTi为一月中第i天的日较差,M为当月天数。按照同样的方法计算可得土壤含水量的相关数值。我们在分析土壤温度和含水量的年内季节动态时采用5年数据的平均值,因为我们更关注季节变化而非年际差异。采用成对样本T检验对比不同监测点的热量指标和生长季指标均值差异。
为了便于与全球林线温度相比较,本文按照Körner 和 Paulsen的方法[24],利用10cm土壤温度定义芦芽山林线处树木的生长季长度:春季土壤日均温首次达到3.2℃为生长季的开始,秋季土壤日均温首次低于3.2℃为生长季结束,生长季长度为开始日和结束日之间的天数。并且,定义土壤日均温开始连续5天小于0℃为土壤冻结开始,以5天中的第一天为冻结开始日,土壤日均温开始连续5天大于0℃为土壤解冻开始,以5天中的第一天为解冻开始日[19, 35]。
2 结果
2.1 芦芽山针叶林分布上限土壤温度的季节变化
芦芽山南、北坡林线土壤年均温分别为-0.54℃和-0.83℃,二者存在显著差异(t=1.97,P< 0.01)。整体上,南、北坡林线土壤温度先上升后下降呈近似余弦函数曲线变化。北坡林线土壤最冷月为2月,南坡土壤最冷月则为1月,南北坡土壤最热月出现的时间一致,都为8月。北坡林线土壤在10月底冻结,解冻日则出现在5月上旬,土壤冻结时间平均长达192d。南坡林线土壤冻结日滞后于北坡6d,出现在11月上旬,解冻日同样滞后于北坡出现在5月中旬,因此南坡土壤冻结时长与北坡大致相同(图3)。北坡林线土壤日较差在11月最小,在6月最大。而南坡林线土壤日较差的最小和最大值则分别出现在4月和6月。除5月、9月、10月和11月之外,北坡林线土壤日较差的月均值都在南坡之上(图4)。
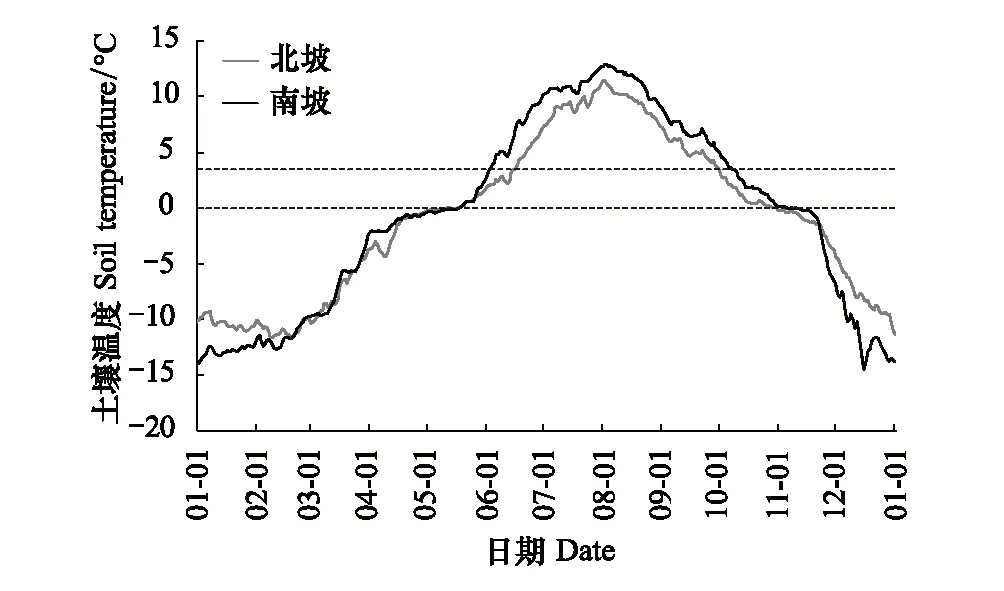
图3 芦芽山针叶林分布上限南北坡土壤温度的季节变化 Fig.3 Seasonal changes of soil temperature at southern and northern treeline on Luya Mountain
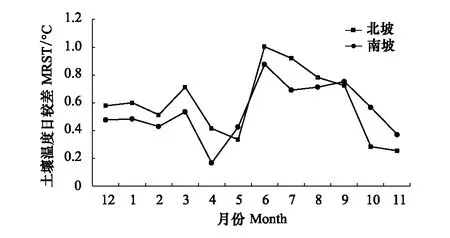
图4 芦芽山针叶林分布上限南北坡林线土壤温度日较差(MRST)季节变化Fig.4 Variation of monthly mean of soil temperature diurnal range (MRST) at southern and northern treeline on Luya Mountain
2.2 芦芽山针叶林分布上限土壤含水量的季节波动
年际间,芦芽山林线土壤含水量呈现循环式变化。冬季和早春(11月至来年4月)土壤含水量低且变化稳定,春季末随着土壤解冻和冰雪融化土壤含水量(5月)急剧增加,夏季和秋季中期(6月至10月)土壤含水量较高但波动也较大,秋季末(11月)因土壤冻结,含水量急剧减小直到再一次达到稳定状态,一个循环完成。土壤含水量在冻结日之前开始回升,在解冻日后仍然保持下降。南北坡林线土壤均在土壤完全解冻后(5月末)、夏季和秋季中期(7月中旬及9月底)出现波峰,而在8月中旬土壤含水量降低明显。另外,南坡林线未冻水含量大于北坡(图5)。
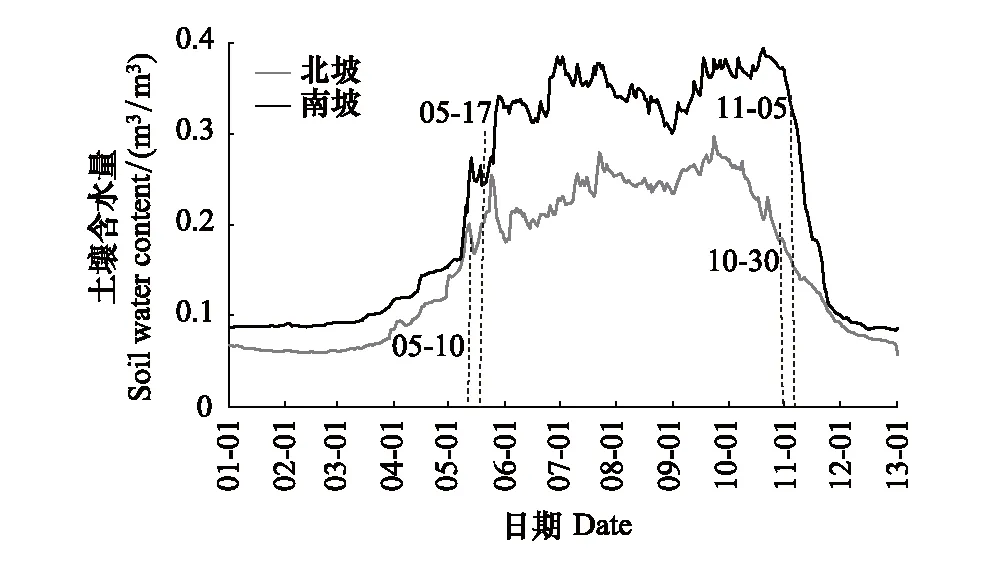
图5 芦芽山南北坡林线土壤含水量(SWC)的季节动态Fig.5 Seasonal dynamics of soil water content (SWC) at southern and northern treeline on Luya Mountain
2.3 基于土壤温度的针叶林分布上下限生长季估算
基于土壤温度估算的生长季参数显示,芦芽山北坡林线生长季开始于6月中旬结束于9月底,生长季长度为110d左右。南坡林线生长季开始提前于北坡约一周左右,结束日期延后约5d,南坡林线处针叶林生长季长度相对于北坡延长约12d,但是南北坡林线处生长季开始、生长季结束和生长季长度均没有显著性的差异。而在针叶林分布下限处(2040m),生长季开始显著早于林线处,并且生长季结束推迟显著,因此生长季长度显著大于林线处(表1)。
2.4 针叶林分布上下限土壤温度和含水量对比
芦芽山针叶林分布下限(海拔2040m处)年内温度变化格局与林线处大致相同。具体的,海拔2040m处土壤年均温为3.18℃,显著大于林线处年均温(t=1.90,P< 0.001),土壤最热月与林线处一致,出现在8月,最冷月(2月)只与北坡林线相同。芦芽山针叶林分布下限土壤解冻日分别提前于南北坡林线土壤12d和5d,冻结日分别延后34d和40d,因此土壤解冻时长达219d,冻结时长仅为147d,并且在冬季和早春冻结状态下土壤温度比林线土壤更稳定,并未出现土壤极端低温(图6,表2)。

表1 芦芽山针叶林上下限生长季参数估算
GSL表示生长季长度GSL denotes growing season length;表中不同字母代表均值差异显著
芦芽山针叶林分布下限处生长季内土壤含水量为0.223m3/m3,显著低于南北坡林线土壤含水量。并且,土壤含水量最高值出现在6月,然后逐月降低,而南北坡林线处土壤含水量最高值分别出现在10月和9月(图6,表2)。这可能是因为针叶林分布下限土壤水分主要来源于上一年的融雪水,而林线土壤水分则受秋季降水影响较大。
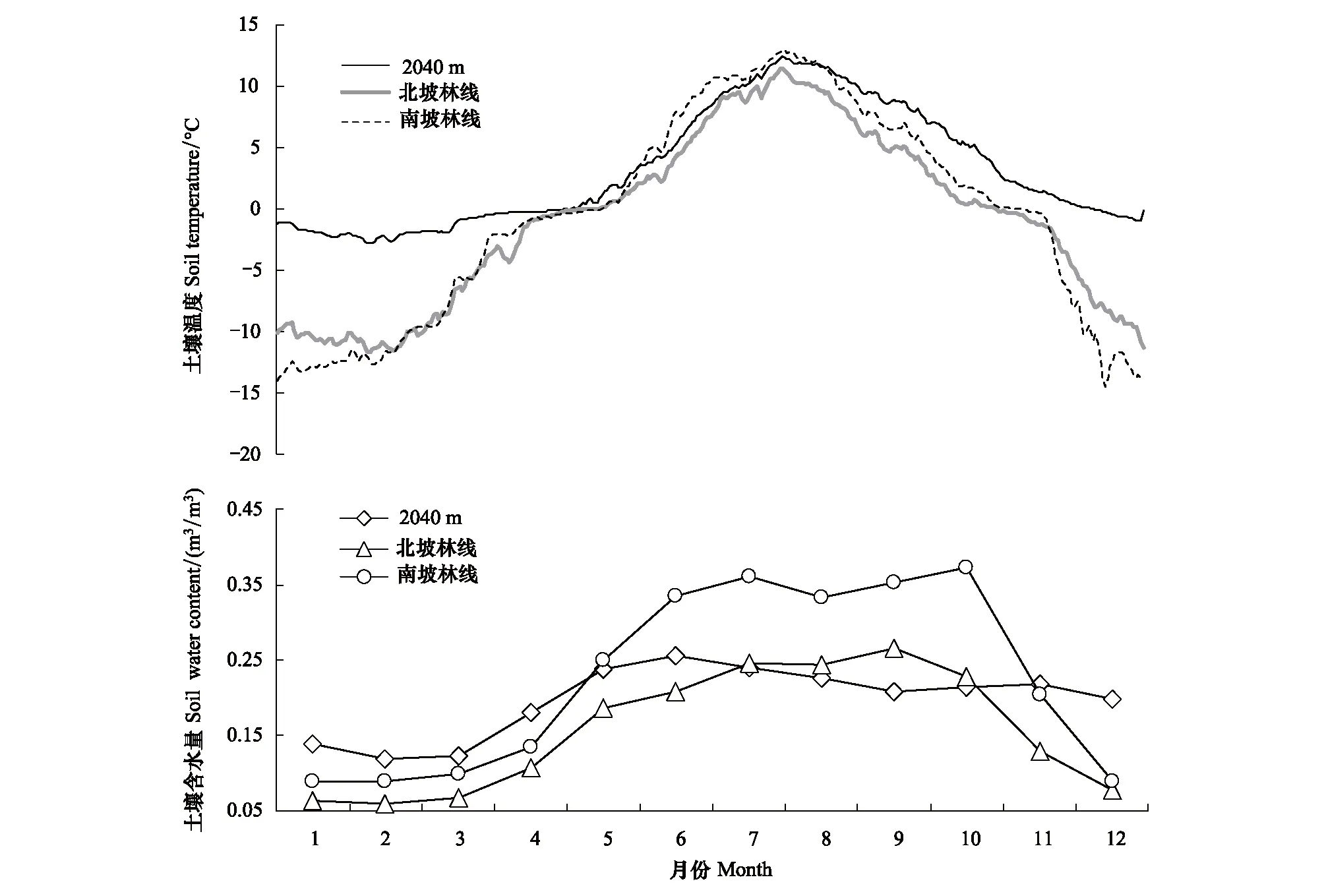
图6 芦芽山针叶林分布上下限土壤温度(ST)和含水量(SWC)的对比Fig.6 Comparison of soil temperature (ST) and water content (SWC) at upper and lower limits of coniferous forest on Luya Mountain
表2 芦芽山针叶林分布上下限土壤水热指标的对比
Table 2 Comparison of soil temperature and moisture indices at upper and lower limits of coniferous forest on Luya Mountain

土壤年均温AMST/℃生长季土壤均温MST/℃土壤最热月均温WMST/℃生长季土壤含水量MSWC/(m3/m3)2040 m3.18±0.46a9.17±0.51a12.41±0.47a0.22±0.01a北坡林线Treeline at north slope-0.83±0.42b7.60±0.42b9.64±0.92b0.25±0.01b南坡林线Treeline at south slope-0.54±0.30c8.12±0.50b11.52±0.55c0.35±0.03c
AMST表示土壤年均温Annual mean of soil temperature; MST表示生长季土壤均温Mean soil temperature in growing season;WMST表示土壤最热月均温Mean of soil temperature in the warmest month;MSWC表示生长季土壤含水量 Mean soil water content in growing season
3 讨论
3.1 芦芽山土壤温度和含水量季节变化及二者的耦合关系
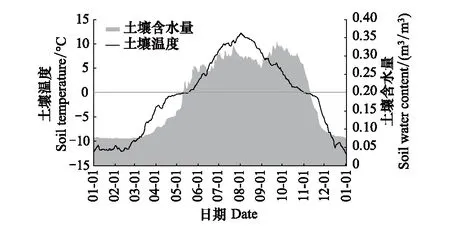
图7 芦芽山土壤温度(ST)和含水量(SWC)的季节变化Fig.7 Seasonal changes of soil temperature (ST) and water content (SWC) on Luya Mountain
本研究结果显示,芦芽山土壤温度和含水量随季节变化都是先升高后下降,展现出水热同季的土壤气候条件。从季节角度看,春季初,芦芽山林线土壤水分在土壤解冻日之前开始回升,并且这个时间段内土壤温度也快速回升(图7),这与念青唐古拉山林线土壤温湿度变化的研究结果相符[19]。这可能主要是因为,春季初白天太阳辐射增强而导致积雪缓慢融化,地表水分含量有所提升,而在夜间辐射冷却使得融水(或潮湿的地表)再冻结,而这个过程中释放的凝结潜热也将进一步导致土壤升温。陈海存等的研究也表明在春季冻结日之前土壤温度和含水量相关系数可达0.8[36]。另外,我们还发现,当土壤含水量回升到0.2 m3/m3时,土壤在0℃左右保持了长达7d的稳定时期。在解冻日附近,地表吸收的太阳辐射热量主要用于完成水分的相变,土壤升温减缓[37]。同理,在冻结日左右,土壤水分冻结释放的凝结潜热也导致芦芽山林线土壤秋季降温幅度减小。在夏季和秋季初的土壤完全解冻期,本研究发现土壤含水量在土壤最热月(8月)达到最低,而含水量高值出现在土壤降温迅速的秋季中期。这其中除了降雨季节变化的影响外,土壤高温而导致的土壤水分蒸散也同样不可忽视。因此,芦芽山林线树木生长,尤其是在干旱年,可能会在夏季受到干旱胁迫。
土壤温度日较差是反映土壤水热关系的主要指标之一。由于水的热容量远大于土壤土体本身,土壤水分含量和状态成为影响土壤温度日较差的主导因素[16, 18]。本文的研究结果也证实了这一点。我们发现,在土壤逐渐解冻和冻结的月份,土壤温度日较差达到全年最低,同时,在土壤完全解冻的夏季,土壤含水量高的南坡土壤日较差小于北坡。
3.2 芦芽山针叶林分布上限不同坡向土壤温湿度对比
一般说来,由于太阳辐射的不均匀分配,南坡土壤温度高于北坡,而土壤含水量低于北坡。本研究中,芦芽山南坡林线土壤年均温(-0.54℃)显著大于北坡(-0.83℃),支持了这一结论。然而,从季节角度看,研究结果显示秋季末和冬季芦芽山南坡林线土壤温度小于北坡。这可能是由于秋冬季节北坡较厚雪被的保温作用所导致。山地小气候复杂多变,而就降水来说,迎风坡多于背风坡,但降雪量,特别是积雪深度,往往是背风坡高于迎风坡[22]。季节性雪被具有较强的绝热能力和低热传导率特性,可以有效地减缓土壤与空气之间的热量交换并且阻隔土壤热量的散失[38-39]。雪被的阻尼效应与其自身厚度有关,较厚雪被因热容量大而对温度变化的敏感性更低[40]。我们还发现芦芽山林线处,秋季降雪初期,积雪深度更大的北坡土壤均温开始大于南坡,并且日较差开始小于南坡。相对的,无雪期北坡土壤日均温小于南坡,日较差大于南坡。
与一般结论不同,芦芽山南坡林线土壤含水量显著大于北坡。土壤含水量的变化不仅受光照强度的影响,还受降水量和植被的影响[41]。芦芽山地区北部的朔州气象站(39°18′N,112°26′E)记录的年均降水量为402.2mm,低于南部的五寨气象站(459.8mm)。并且,与草地和草甸不同,森林因冠层郁闭、枯落物覆盖而具有更好的水源涵养能力。因此,芦芽山降水量较大的阳坡土壤含水量更高。这与佘波和武晓红的研究结果相符。他们通过对比发现,太原东山试验林场裸地土壤含水率总体表现为阴坡>半阴坡>阳坡,而林地土壤含水率总体表现为阳坡>半阳坡>阴坡[42]。高山林线是山地郁闭森林与高山灌丛或草甸之间的生态过渡带,这一交错区的边界受到诸多驱动因素的影响[9,24]。 因此,在实地设置监测点位时,难以确保南北监测点到林线边界距离以及森林郁闭度完全一致,这也使得本研究存在一定的局限性,可能导致研究结果受到其他局地因子的影响,比如,杨艳刚等对芦芽山草甸土壤含水量的研究,发现由于坡度的原因,上坡位补水导致阳坡靠近林线的下坡位草甸土壤含水量增大[43]。
3.3 芦芽山针叶林分布上下限土壤温度及含水量的差异
基于多年的实测数据,本研究计算对比芦芽山南、北坡林线生长季土壤均温、生长季长度、土壤最热月均温和生长季土壤含水量发现,除生长季土壤均温(南坡8.1℃,北坡7.6℃)外,南、北坡林线其他土壤热量指标均表现出显著差异(表2)。这一结果说明,生长季土壤均温可以更好地指示林线海拔所处的高度,且与中国其他地区以及全球相关研究的结论均一致[24, 28]。具体的,芦芽山北坡林线土壤生长季均温为7.6℃,符合全球暖温带林线生长季土壤均温(7—8℃),进一步说明了热量不足对北坡林线树木生长的限制。然而,南坡生长季土壤均温(8.1℃)大于全球均值,与Müller等在喜马拉雅高山林线的研究结果一致[44]。更高的土壤温度和含水量,说明芦芽山南坡林线树木生长可能还受到其他局地因子的影响。我们还发现,相较于北坡林线,南坡林线秋冬季节有更低的土壤温度。更低的土壤温度致使南坡冬季冻土层深厚,土壤春季解冻缓慢可能会抑制树木茎干的水分恢复和生长启动,进而影响树木生长[45-46]。
本研究通过对比高海拔林线与低海拔森林分布下限(2040m)土壤水热指标及其季节变化,我们发现芦芽山高海拔林线土壤热量指标全部显著小于低海拔,而生长季土壤含水量却显著大于低海拔。这一山地气候特征多见于我国干旱和半干旱区,并对海拔梯度上生长-气候关系产生重要影响。尽管本研究并未涉及树木径向生长的同步监测,但在同一地区,研究发现芦芽山低海拔华北落叶松树轮年表与生长季的降水因子呈正相关,而高海拔处均与生长季中降水因子呈负相关,并且认为低海拔树木生长受干旱限制而高海拔受低温限制[47-48]。这进一步证实了芦芽山高海拔冷湿而低海拔暖干的环境条件。另外,我们还发现芦芽山针叶林分布上下限土壤含水量有不同的季节变化特征,针叶林分布下限处土壤含水量最高值出现在土壤完全解冻后的6月,而南北坡林线处土壤含水量最高值分别出现在10月和9月的秋雨季。这一结果从侧面也得到了张文涛等的研究结果的支持。他们发现芦芽山低海拔树轮差值年表与当年夏季降水量呈极显著正相关,并且与6月份的温度呈负相关,高海拔年表与降水具有极显著负相关关系,与当年5月平均气温存在显著正相关关系[49]。结合本研究和张文涛等的结果可以看出,即便是在芦芽山针叶林分布下限暖干的环境条件下,土壤含水量最大的6月份,水分仍然是芦芽山针叶林生长的限制因子,而在冷湿的高海拔林线区,降水的限制作用已然消失,针叶林生长更多受到生长季内温度的影响。
4 结论
本研究发现芦芽山土壤温度和含水量随季节变化先升高后下降,土壤温度与土壤水分含量和状态之间的耦合关系影响着二者的季节变化。芦芽山针叶林分布上限南坡土壤年均温和含水量显著大于北坡,而南北坡生长季内土壤均温无显著差异。在针叶林分布下限,土壤热量指标均明显高于林线处,而生长季土壤含水量却显著低于林线处。并且,针叶林分布下限土壤水分含量在春季融雪结束后的6月达到最大,而南北坡林线处土壤含水量的最大值却出现在秋雨季(10月和9月)。研究结果证实了芦芽山高海拔土壤冷湿而低海拔暖干的环境条件。本研究也存在一定的局限性,今后的工作可以将树木茎干生长动态纳入到监测之中,设置更多土壤深度梯度及海拔梯度的监测点位,并以本文的结果为基础继续深入研究华北地区山地土壤温湿度特征及差异。
