羌塘高原降水梯度植物叶片、根系性状变异和生态适应对策
2020-03-12赵广帅石培礼张宪洲
赵广帅,刘 珉,石培礼,宗 宁,张 鑫,张宪洲,3
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 2 国家林业和草原局经济发展研究中心, 北京 100714 3 中国科学院大学资源与环境学院, 北京 100190
在长期进化过程中,植物为了抵御各种环境压力而逐渐形成了许多自身生存和生长发育的响应与适应策略,这些策略具体表现为植物形态和生理特征(即植物性状)的变化[1],其中对环境变化产生响应或对生态系统功能产生影响的植物性状称为植物功能性状[2- 3]。植物功能性状是连接植物与环境的桥梁,阐明不同器官性状和功能的关系是理解植物演化的关键[4],特别是不同地理分区或分类类群叶片、根系性状对比研究,对于理解植物性状的进化和表达,资源的获取、物种分布以及生态系统对干扰的响应具有重要作用[5- 8],也将极大的改进全球生态系统功能模型和气候变化模型的可预测性[9- 11]。
尽管植物功能性状研究已被广泛关注,但不同尺度植物性状间关系及其对环境响应的量化研究仍然较少[12- 13]。目前,大多数研究主要集中在局地站点尺度[5,14],然而更大尺度上不同气候、土壤和地形等因素对植物功能性状影响尚不明确[15- 16],特别是对高寒干旱环境梯度带的植物功能性状变化研究更少[17]。此外,大部分研究集中于地上叶片性状的研究,主要分析性状间简单的回归关系[13, 18],而对地下根系性状研究较少,尤其是地上、地下成对性状关系的研究[19- 22]。目前,青藏高原环境梯度带的植物地上、地下功能性状是协同还是权衡关系的研究还鲜有报道。
植物功能性状是长期进化和环境因子双重作用的结果,与植物资源获取、利用和保持能力紧密联系[23- 24]。通常研究认为,植物不同器官具有一致的性状特征,地上、地下性状主要是协同变化而权衡作用较小[25- 27]。例如,Craine等[28]基于站点的研究认为,不同物种叶片和根系性状表现出一致性变化。Liu等[29]在内蒙古草原环境梯度带上的研究也表明,地上、地下器官权衡变异较小,更多的是协同变化。在环境梯度带上或不同物种间,叶片和根系性状具有相似的表现型,如养分贫瘠地区的植物叶片和根系趋向于低养分含量、高的结构投入和长寿命等[30- 31]。从进化角度来看,资源获取型物种具有高的资源获取能力、高的组织养分含量、比叶面积(SLA)和比根长(SRL)以及净光和速率等;而资源保持型物种具有高的资源保持能力、低的组织养分含量、高的养分回收效率和抗压能力等[32- 34]。然而,环境因子能够限制或改变性状间关系的进化或表达形式[22]。
植物功能性状深受土壤养分和气候因子的影响[35- 36],特别是环境梯度上植物功能性状的变异反映了植物各类适应机制的相对重要性以及与气候、土壤、地形等驱动因子的交互作用[37- 38]。Ordoez等[13]发现在不同的土壤养分条件下,植物生长和资源保持性状间存在权衡作用,并且降水对权衡作用影响较大。Craine等[22]在不同草地的研究表明,不同环境条件下叶片和根系性状间关系并不一致。养分限制可能改变根系而对叶片构建基本无影响,植物具有一套低氮适应的叶性状未必具有完整的低氮吸收的根系性状。此外,地形等因素也会对植物性状产生影响[39]。Craine等[19]在新西兰30个草原站点的研究表明,不同海拔高度植物性状存在显著差异。尽管叶片和根系性状的关系已有较多研究,然而在不同区域二者关系是表现为协同还是权衡,并且气候、资源有效性、胁迫和干扰是否会对其产生影响仍然知之甚少。
作为西藏高原的主体,藏北羌塘高原平均海拔在4500 m以上,气候寒冷干燥,太阳辐射强烈,土壤贫瘠,冻土广泛分布,是生态环境最为脆弱的区域[40]。羌塘高原由东向西存在明显降水梯度带,年降雨量(MAP)从最东部700 mm左右降低到最西端的50—80 mm左右。在此环境梯度上土壤有机质从4.0%左右降低到1.0%以下,土壤全氮(TN)含量从0.2%降低到0.02%左右[41],水分及与其密切相关的养分梯度对高寒草原物种分布和群落结构产生更重要影响。因此,羌塘高原降水梯度带植物地上、地下功能性状必然存在与高寒、干旱、贫瘠环境相适应的特征,而且地上和地下器官可能在碳、氮和水获取之间存在协同或权衡,我们选择3组具有代表性的叶片和根系成对性状[13, 27, 29],重点阐明受长期进化和环境因子双重作用,羌塘高原降水梯度带不同物种地上、地下成对性状是如何变化的?与光合作用有关的叶片性状和与水分、养分吸收相关的根系功能性状在区域变化是否存在相互联系?是协同还是权衡关系?功能性状的变化是如何适应环境变化的?
1 材料与方法
1.1 研究区概述
羌塘高原位于西藏自治区北部,南起冈底斯—念青唐古拉山脉,北抵昆仑山脉,东迄青藏公路西侧91°E左右的内、外流水系分水岭,西止于国境线,面积约60万km2,行政区划上属西藏自治区的那曲与阿里两地区管辖。羌塘高原是我国气候条件最为恶劣、生态环境极为脆弱的典型区域,环境特点主要表现为气候寒冷、干旱、多风,地表沙物质丰富,植物稀疏低矮等,生态容量较低。羌塘高原干旱气候特征显著,蒸发强度普遍>1800 mm,年均风速多在3 m/s以上,年平均干燥度指数1.6—20[42]。羌塘高原气候寒冷,大部分地区年平均气温< 0 ℃,最冷月(1月)均温为-10—-18 ℃,而最暖月(7月)均温大多不及10 ℃,全年> 0 ℃的活动积温多在1200 ℃以下[43]。羌塘高原面的年均温变化不超过2C,但羌塘高原东西端降水量梯度变化显著,相差600mm左右[44]。
1.2 数据获取
1.2.1植物性状取样和指标分析
在2014年8月生长季高峰期,沿羌塘高原降水梯度带(那曲县罗玛镇至改则县)共选择13个站点进行草地群落优势种叶片、根系和土壤等样品采集(表1)。为了减小地形和放牧的干扰,所有站点都位于地势平坦,远离人类栖居地,植物生长良好的地带性植被区域,相邻站点间隔50—80 km,确保取样点的代表性。每一站点选取全部优势物种,即代表80%—90%草地生态系统生物量的物种[45],每一物种最少选取20株生长良好的个体采集完全展开的绿叶,至少5株采集根系样品。同时,在每个站点随机采集3个土壤剖面,土壤剖面间距离500 m以上,每个土壤剖面采集0—20 cm土层的土壤样品。

扫描的叶片和根系图像分别使用Adobe Photoshop CS 5软件和WinRHIZO 软件(Regent Instruments Inc., Quebec City, QC, Canada),获取叶面积和根长度,计算叶片和根系性状相关指标:
SLA=Larea/Lmass
(1)
SRL=Rlength/Rmass
(2)
LNarea=LNmass×Lmass/Larea
(3)
RNlength=RNmass×Rmass/Rlength
(4)
式中,SLA(cm2/g)为比叶面积,SRL(cm/g)为比根长,Larea(cm2)和Lmass(mg)分别为扫描图像叶片总面积和干重,Rlength(cm)和Rmass(mg)分别为扫描图像根系总长度和干重,LNarea(mg/cm2)和LNmass(mg/g)分别为叶片单位面积氮含量和单位质量氮含量,RNlength(mg/cm)和RNmass(mg/g)分别为根系单位长度氮含量和单位质量氮含量。
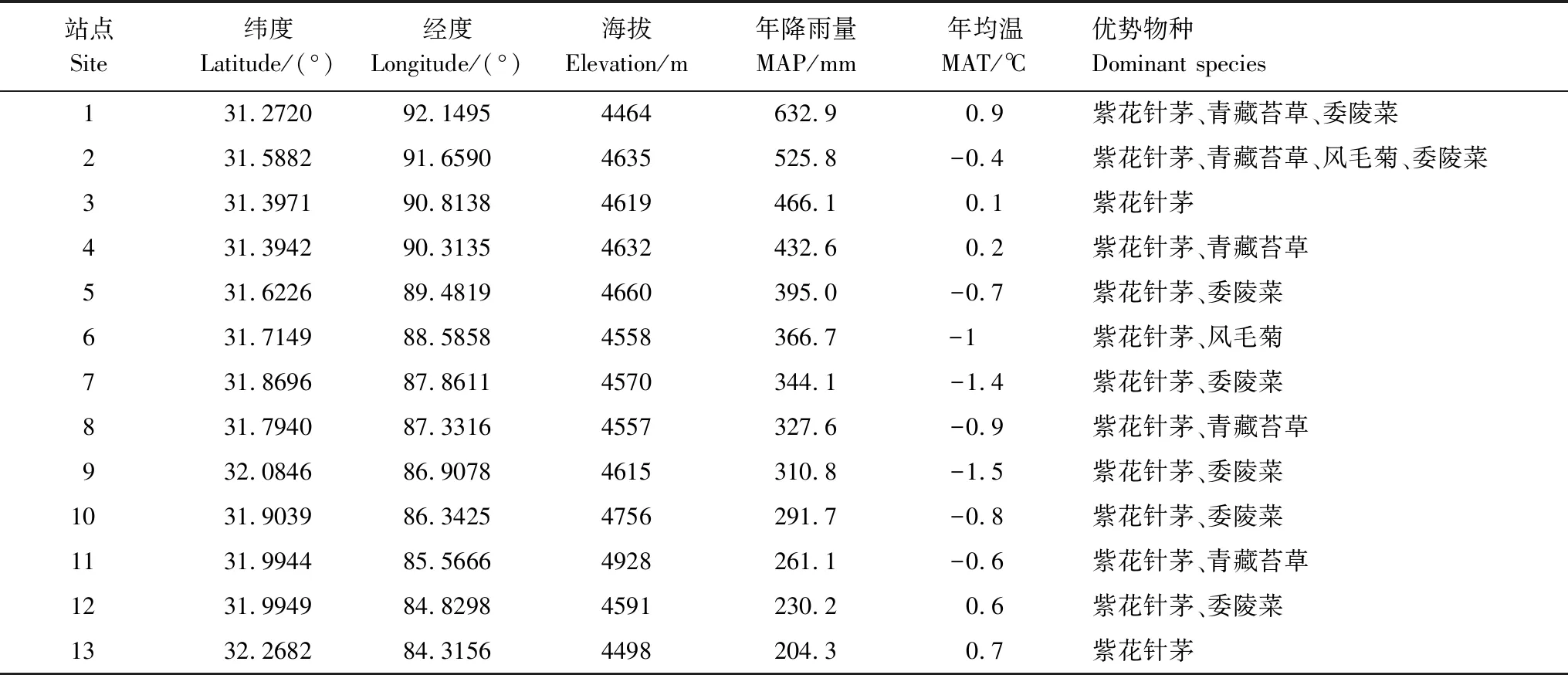
表1 采样点位置及环境特点
MAP: mean annual precipitation; MAT: mean annual temperature
1.2.2气象数据
年均降雨量(MAP,mm)和年均温(MAT,℃)数据来自国家气象站(6个)和中国科学院拉萨高原生态综合试验站在藏北羌塘高原设置的HOBO自动气象观测站(11个)。每年的潜在蒸发量(PET)主要根据气象观测数据,利用cropwat 8.0软件计算获得(http://www.fao.org/nr/water/infores _databases_cropwat.html),地面太阳辐射数据(Ra,kWh m-2d-1)主要来自NASA提供的地面气象和太阳能数据集(https://eosweb.larc. nasa.gov/cgi-bin/sse/grid.cgi)。
1.3 数据处理和分析
采用基于类型Ⅰ平方和分解来量化不同变异组分的嵌入式方差分析法(Nested ANOVAs)[29]区分不同站点和物种间叶片和根系性状的变异性(Nested Procedure, SAS version 9.1; SAS Institute Inc., Cary, NC, USA)。
羌塘高原降水梯度带叶片和根系成对性状(SLA-SRL,LNmass-RNmass,LNarea-RNlength)间异速生长关系分析方法如下:先对各性状值进行对数转换,使其满足正态分布;然后采用标准主轴回归(Standardized Major Axis,SMA)计算成对性状异速生长方程的斜率、截距和性状间的相关系数;通过计算残差和拟合斜率之间的相关性,检验拟合斜率与假定斜率1的差异性[46]。以上分析均采用基于DOS系统的SMATR软件包[47]进行。因为羌塘高原植物地上和地下性状主要是由于降水差异和分类群变化引起的,所以进一步分析不同降水区间和植物分类群对地上和地下成对功能性状的影响。
采用双变量线性混合模型分析植物性状与土壤养分和气候的关系。所有数据先进行正态分布检验(单样本K-S检验),土壤养分和降水数据进行对数转换,以满足正态分布,模型采用最大似然估计法(maximum likelihood methods)。由于不同站点间植物性状可能存在差异,将土壤和气候及其交互影响作为固定效应,而不同站点作为随机效应。上述分析全部在SPSS 19软件(SPSS Inc., Chicago, IL, USA)中进行。
2 结果分析
2.1 取样站点和物种的地上、地下功能性状的变异

图1 站点和物种对地上和地下性状变异的解释度Fig.1 Variance components of leaf and root traits across sites and speciesSLA:比叶面积specific leaf area;LNmass:单位质量叶氮含量unit mass leaf nitrogen content;LNarea:单位面积叶氮含量leaf nitrogen content per unit leaf area;SRL:比根长specific root length;RNmass:单位质量根氮含量unit mass root nitrogen content;RNlength:单位长度根氮含量root nitrogen content per unit root length
分析不同站点以及同一站点不同物种间叶片和根系性状的变异特征,结果表明除同一站点不同物种间LNmass变异不显著外(P= 0.57),不同站点或不同物种间叶片和根系性状都存在显著变化(P< 0.05),但二者对于不同性状的解释量不同。不同站点对于叶片性状SLA、LNmass、LNarea解释量分别为43%、92%、69%,而不同物种对于根系性状SRL、RNmass、RNlength解释量分别为77%、31%、92%(图1),整体来看,不同站点间叶片性状变异较根系性状变异大,而不同物种间根系性状变异更大。
2.2 地上、地下植物性状沿环境梯度的变化特征
将样带上植物综合角度,分析了羌塘高原环境梯度带上气候因子和土壤因子对叶片和根系性状的影响。在气候因子中,对地上和地下性状影响最重要的因子是年降水量的变化,随着MAP增加,SLA和SRL分别呈轻微显著增加和降低趋势,而LNmass、RNmass和LNarea呈显著降低趋势。随着Ra升高,SLA轻微显著降低,SRL、LNmass轻微显著增加,只有RNmass显著增加。MAT和PET基本没有影响(图2)。
对地上和地下性状影响最主要的土壤因子是土壤C∶N,其他土壤养分因子和海拔基本没有影响。随C∶N比增加,LNmass、RNmass和LNarea都呈显著增加的趋势,土壤TN只对RNmass造成显著影响(图3)。

图2 植物地上和地下性状与气象因子的关系Fig.2 The relationships between leaf/root traits and climatic factors 显著线性相关用实线图示,而微弱相关用虚线图示;MAP:年降雨量mean annual precipitation;MAT:年均温mean annual temperature;PET:潜在蒸散发potential evapotranspiration;Ra:辐射total daily radiation;SLA:比叶面积specific leaf area;LNmass:单位质量叶氮含量unit mass leaf nitrogen content;LNarea:单位面积叶氮含量leaf nitrogen content per unit leaf area;SRL:比根长specific root length;RNmass:单位质量根氮含量unit mass root nitrogen content;RNlength:单位长度根氮含量root nitrogen content per unit root length

图3 植物地上和地下性状与土壤养分和海拔的关系Fig.3 The relationships between leaf/root traits and soil nutrient and elevation 显著线性相关用实线图示,而微弱相关用虚线图示;TN:土壤总氮soil total nitrogen content;C∶N:土壤碳氮比soil carbon and nitrogen ratio;IN:土壤无机氮inorganic nitrogen;E:海拔elevation;SLA:比叶面积specific leaf area;LNmass:单位质量叶氮含量unit mass leaf nitrogen content;LNarea:单位面积叶氮含量leaf nitrogen content per unit leaf area;SRL:比根长specific root length;RNmass:单位质量根氮含量unit mass root nitrogen content;RNlength:单位长度根氮含量root nitrogen content per unit root length
从不同功能群植物来看,苔草类(青藏苔草)和禾草类(紫花针茅)的植物地上和地下性状随降水梯度呈现显著的变化趋势,其中青藏苔草最为明显,除了LNmass外,其他性状如所有物种放在一起一样随降水都呈显著的变化规律;紫花针茅除了SLA和SRL外,其他四个性状都随降水梯度呈现有规律的变化(图4)。

图4 不同分类群植物性状随降水梯度的变化Fig.4 The variation of leaf/root traits in different taxa along with precipitation gradient显著线性相关用实线图示,而微弱相关用虚线图示;P. b.:二列委陵菜Potentilla bifurca;C.m.:青藏苔草Carex moocroftii;S.p.:紫花针茅Stipa purpurea;S. j:风毛菊Saussurea japonica;SLA:比叶面积specific leaf area;LNmass:单位质量叶氮含量unit mass leaf nitrogen content;LNarea:单位面积叶氮含量leaf nitrogen content per unit leaf area;SRL:比根长specific root length;RNmass:单位质量根氮含量unit mass root nitrogen content;RNlength:单位长度根氮含量root nitrogen content per unit root length
2.3 地上和地下成对功能性状的关系
羌塘高原地上、地下成对性状(SLA-SRL,LNmass-RNmass,LNarea-RNlength)间呈显著相关性,其中SLA和SRL、LNarea和RNlength呈极显著负相关(表2)。尽管LNmass与RNmass呈极显著正相关,但异速生长斜率显著小于1,表明二者间存在着显著的异速生长关系(表2)。
不同降雨区间或不同物种间地上、地下成对性状异速生长关系存在显著差异,其中不同降雨区间或不同物种间LNmass-RNmass异速生长斜率存在极显著差异;尽管SLA-SRL、LNarea-RNlength拥有相同的异速生长斜率,但截距存在显著差异(表3)。具体来看,相对干旱端(<400 mm)和湿润端(>600 mm)的SLA-SRL、LNmass-RNmass和LNarea-RNlength成对性状仍保持显著负相关,而中间区域(400—600 mm)基本呈正相关关系(表3),说明干旱端和湿润端地上、地下成对性状之间具有更强烈的地上与地下权衡策略,中间降水区域为协同关系。与其他物种相比,青藏苔草的SLA和SRL、LNarea和RNlength基本都是呈负相关关系,异速生长斜率为负值;而且青藏苔草和紫花针茅LNmass和RNmass异速生长斜率明显小于1(表3),说明这两个优势植物的地上、地下成对性状之间具有更强烈的权衡策略。
3 讨论
3.1 羌塘高原降水梯度带植物地上、地下性状变异与适应对策
羌塘高原草地样带站点和同一站点不同分类群的植物功能性状方差分析表明,地上、地下性状存在显著变异,不同站点间叶片性状变异较根系性状变异大,而不同物种间根系性状变异更大。说明叶片在站点内相同环境下表现出趋同适应,而在环境梯度上表现出强烈的分异;不同物种的根系性状的种间差异可能更多地决定于植物的系统发育(phylogentic)。植物功能性状的变异主要取决于植物进化差异和环境因子限制[31]。不同物种间性状的差异是进化过程中主要植物谱系间性状传递给后代类群过程中产生分歧的结果(进化趋异),而不同站点间性状的差异是物种对不同环境的塑性响应(适应性进化)[23]。不同物种间性状的显著差异反映了物种进化史的不同,而不同站点间的差异反映了环境因子的影响。
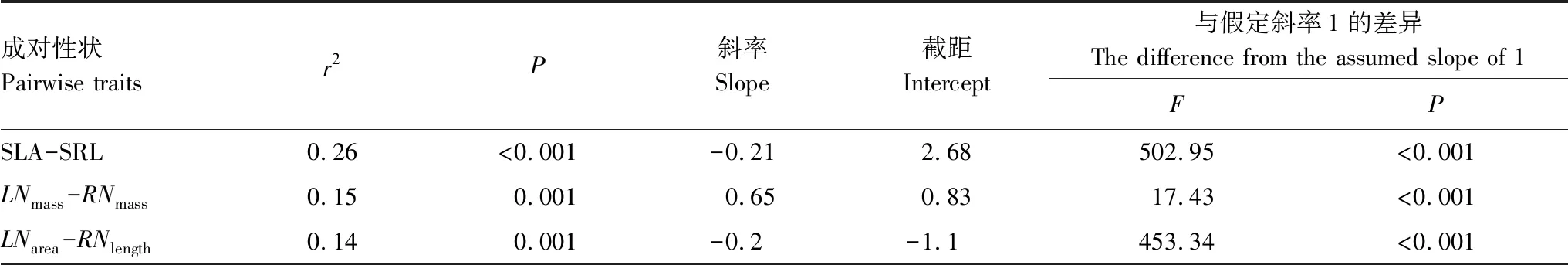
表2 羌塘高原地上地下成对性状的标准主轴回归结果
SLA:比叶面积specific leaf area;SRL:比根长specific root length;LNmass:单位质量叶氮含量unit mass leaf nitrogen content;RNmass:单位质量根氮含量unit mass root nitrogen content;LNarea:单位面积叶氮含量leaf nitrogen content per unit leaf area;RNlength:单位长度根氮含量root nitrogen content per unit root length
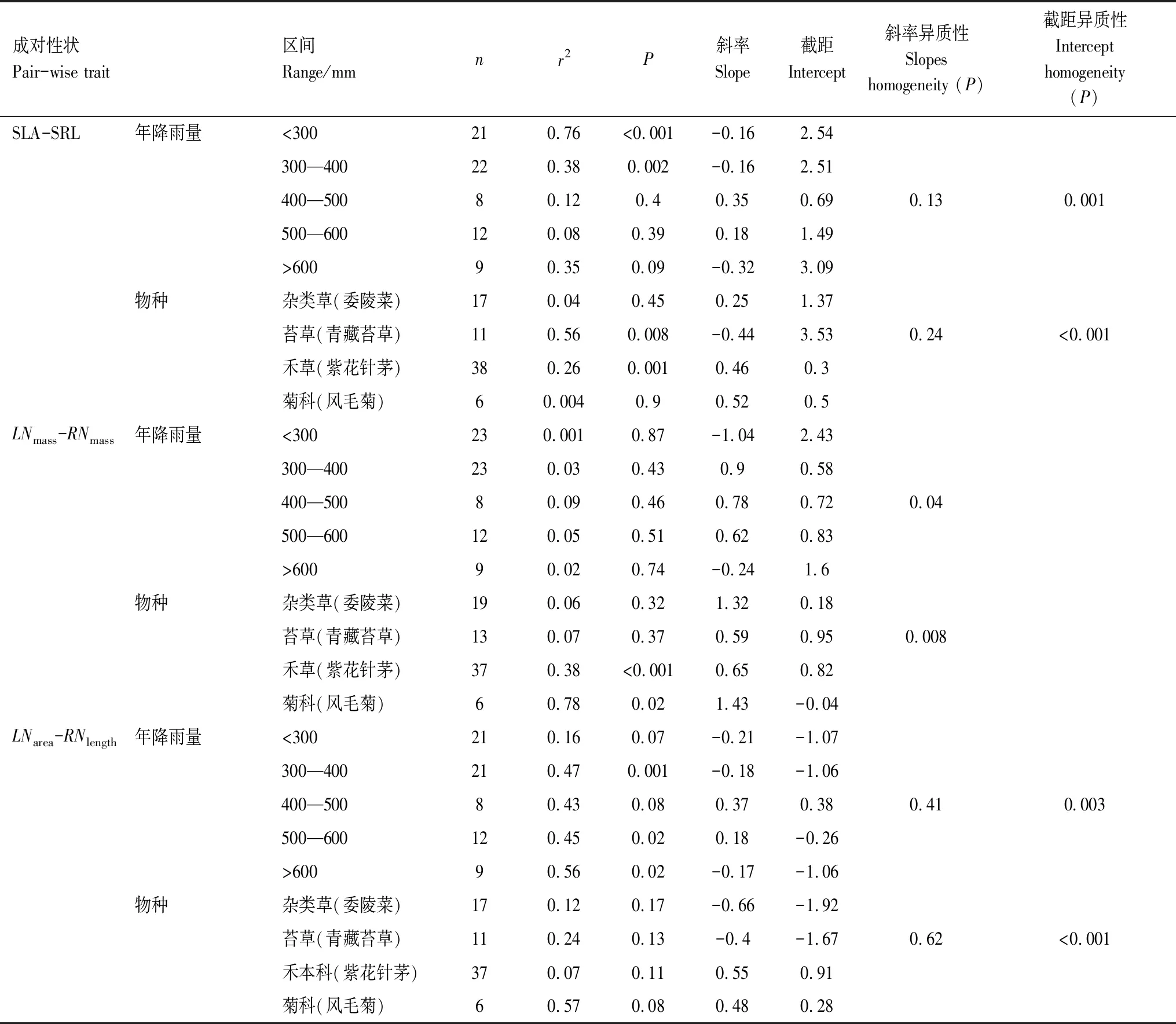
表3 不同降雨区间和植物类群对地上和地下成对性状的标准主轴回归分析
许多研究表明环境因子能够解释大部分植物功能性状的变异[48-50]。羌塘高原降水梯度带不同站点间地上、地下功能性状的显著差异反映了植物地上、地下性状对不同水分及其导致的养分条件变化的适应机制。种间差异是性状变异的重要因素,种间差异解释大量性状变异,这与许多研究一致[16, 27]。Liu等[29]在内蒙古草原的研究结果表明种内和种间差异解释55%以上的地上、地下性状变异。Wright等[26]对全球范围内叶片性状数据的综合分析表明,同一站点内种间差异解释36% SLA变异和38%LNmass变异。物种间的差异主要源于两个方面,一方面是基因决定的稳定性状,不受进化时间和自然选择的影响[23];另一方面,不同物种对同一选择因子或环境条件适应性不同[51]。然而,羌塘高原同一站点不同物种间LNmass差别不大。Hong等[17]在青藏高原的研究数据也表明非豆科植物物种之间LNmass差异不显著。
随降水增加,SLA和SRL分别呈现轻微增加和降低趋势,其中SRL的变化趋势更强一些。表明在西部降水少出现水分胁迫的干旱端植物倾向于利用更高的SRL增加水分和养分吸收能力,而东部降水丰富的湿润端植物倾向于增强光合作用面积提高地上部分的光合生产力。SRL与水和养分吸收紧密相关,被认为是地下竞争能力的形态指标。水分胁迫是生长的限制因素(高寒草原和荒漠草原),植物往往产生长而细的根(较高的SRL)来克服水分胁迫。这种在寒旱条件下高的SRL表明,细根可能更好地装备起来以减缓和适应干旱端土壤水分含量低的胁迫作用。SLA是与光合速率紧密相关的指标,尽管总体随降水增加趋势不显著,但主要优势种青藏苔草的趋势变化非常明显,表明湿润端植物具有较高的光合生产力和地上生物量。这对地上地下的形态性状的变化趋势在先前的研究中也有发现[48],但与一些草地生态系统结果呈现相反的趋势[50]。
辐射对地上地下性状的影响与降水的作用呈相反趋势(图2),这可能与降水高的东部地区湿度大云雾较多,辐射较弱;西部地区降水少,辐射强烈的事实相关。SLA和SRL呈负相关(图2,表2),干旱端植物选择高的SRL和较低的SLA,除了增加根系对水分和养分的吸收外,还可能与干旱区植物降低地上部分生物量投入,减少植物蒸腾作用导致的水分流失有关。
有意思的是,干旱端的植物根系既具有高的SRL,又具有高的叶片和根系的养分含量(LNmass,LNarea和RNmass),这与其他生态系统二者表现不相关[52]甚至呈相反趋势[50]是不一样的,倒是有些木本植物这二者之间有正相关关系[21]。SRL是以根长作为生物量投入的指标,通常反映根系成本和效益的指标。这反映了干旱端植物通过根系单位质量的表面积提高低养分的吸收效率,同时叶片和根系的高养分含量有利于根系的活动和叶片的光合物质生产,保障了根系生长的物质来源。这体现了干旱端植物在地上、地下部分适应低养分和干旱的同时投资适应策略。
从上面比较来看,干旱端植物的地上和地下成对性状具有较强的相对优势。那么,湿润端的高寒草甸植物是不是就没有优势呢?高寒草甸植物又是如何形成比干旱端植物更高的生产力呢?尽管本研究区干旱端叶片的含氮量高于湿润端,但是高寒地区的高海拔植物叶片具有比低海拔植物高的养分含量,比如氮含量甚至比低海拔植物高到4—5倍[53],并且具有相对较高的SLA,优势植物更显著,比如本研究中的青藏苔草在高寒草甸中更明显,这些表明高寒草甸植物还是具有相当优势和有利于植物的光合作用和生产力形成的。再者,湿润端的高寒草甸植物较粗的根系也可以通过丛枝菌根的侵染而提高养分的利用效率[54]。更为重要的是,从降水条件和群落结构来说,高寒草甸具有高的水分条件,群落物种和功能群多样性都相对丰富,叶面积指数高,增加了群落的生产力和稳定性,这是形成高寒草甸高生产力的最重要的因素。
3.2 羌塘高原降水梯度带上地上和地下成对性状的权衡关系
多重的功能性状可能因进化或生物物理限制和权衡等而显著相关[23, 31]。羌塘高原降水梯度带植物的地上和地下成对性状如SLA和SRL、LNarea和RNlength均呈极显著负相关,说明地上和地下获取资源的核心性状存在着显著的权衡关系,即随着植物根系获取水分和养分能力的相对增强,叶片获取光资源和进行光合生产的能力相对减弱。这与以往研究表明随着根系N投入的相对增加,而叶片N投入相对不断减少[55-57]的结果相一致,但与一些叶片和根系协同的研究结果相左[29,52]。尽管本研究发现LNmass和RNmass呈极显著正相关与以往西藏高原的植物的化学计量和全球的种子植物的研究结果一致[58-59],但二者异速生长斜率明显小于1,说明本研究区植物投入根系的养分显著高于叶片,根系获取水分和养分的能力相对增强,而叶片通过光合作用的物质生产能力相对减弱,导致了地上、地下性状间存在极显著异速生长关系,这与羌塘高原高寒草原土壤瘠薄,植物根冠比明显较高的事实相符。
与其他功能群相比,青藏苔草和紫花针茅的地上、地下成对性状存在显著异速生长关系,说明苔草类和禾草类植物地上、地下成对性状之间具有更强烈的权衡策略。在养分贫瘠条件下,莎草科和禾草科植物采取地下部分存储更多养分的权衡策略[60],这可能是莎草科和禾草科植物更适应羌塘高原高寒、干旱环境的原因[61-62]。分布范围相对较窄的伴生物种,其植物组织内部多重生理性状的权衡弱,对环境变异敏感[63-64],而广泛分布的优势种则能够通过多重内部生理性状的权衡来适应外部环境的变化[65-66]。
羌塘高原高寒贫瘠环境会对地上、地下成对性状关系产生显著影响。Liu等[29]在内蒙古草原环境梯度带上的研究表明地上、地下器官权衡变异较小,而更多的是协同变化。然而Geng等[48]在青藏高寒草地与内蒙温性草地对比研究中发现,高寒环境显著改变了地上、地下形态性状间关系。植物地上、地下形态组分不同的环境压力导致叶片与根系性状之间相关性并不一致,高寒低温环境导致青藏高原冻土广布,土壤冻融作用对高寒草甸植物根系限制较大,而对叶片影响较小[65]。尽管羌塘高原降水梯度带LNmass-RNmass呈极显著正相关,但二者存在极显著异速生长关系,这与大多数研究结论并不完全一致[16, 27, 29]。例如,Geng等[48]研究认为无论是不同站点还是不同物种间的LNmass-RNmass表现一致的极显著正相关关系,但地上、地下成对性状间并不存在显著的异速生长关系。然而,Hong等[17]在青藏高原的研究表明不仅不同功能群对土壤养分的响应不同,同一功能群地上、地下部分对土壤养分的响应也不同。地上、地下器官对于资源分配与投入的权衡,将直接决定物种能否在养分贫瘠环境中获得成功[66]。羌塘高原降水梯度带地上、地下成对性状极显著负相关关系或异速生长关系说明,羌塘高原地上、地下性状并不存在一致的协同关系,而是存在强烈的权衡关系。
不同降雨量区间或不同物种地上、地下成对性状的权衡关系更加明显。降水梯度带相对干旱端(< 400 mm的高寒草原和荒漠草原区)和湿润端(> 600 mm的湿润性高寒草甸)地上、地下成对性状呈显著负相关,而中间区域(400—600 mm的高寒草甸)基本呈正相关关系,说明与大多数的高寒草甸区植物的叶片和根系性状协同相比,相对干旱的高寒草原、高寒荒漠和极湿润的高寒草甸的叶片和根系成对性状具有更强的权衡策略。
从羌塘高原从东往西的环境梯度(主要是降水梯度及其导致的土壤养分梯度)上的植物地上和地下性状的权衡关系来看,未呈现叶片和根系性状的经济谱,即没有出现叶片化学性状随降水梯度和养分的协同变化。但是,干旱端叶片和根系较高的单位质量和单位面积的养分含量并不代表植物性状是养分获取型(acquisitive strategy),相反更大程度地体现了养分保持型策略(conservative strategy)。因为前者是以快速获取、低回收的“开源型”养分利用策略;而后者是慢吸收、养分保持和高回收的“节流型”养分利用策略。尽管干旱端植物的养分含量较高,但植物并没有采取快速生长和快速分解的策略,而凋萎叶片具有更高的养分回收效率[44, 67],都体现了干旱端植物的保守性。这与亚高山暗针叶林养分保持型植物叶片保持相对较高的养分含量、较长的叶子寿命和较低生长速率类似。干旱端植物保持较高的养分含量更多地与抵御环境胁迫有关,而与快速生长的获取型策略有显著区别。
3.3 环境因子对成对性状关系的影响
不同生境条件差异,引起植物资源获取和利用策略的选择梯度,是对叶片和根系性状协同或权衡的最适应解释[68],以往许多研究结果差异也是由环境因子(例如气候和土壤因子等)不同造成的[49]。本研究结果表明,不仅不同环境因子对地上、地下成对性状的影响不同,而且同一环境因子对地上、地下成对性状限制也不同。地上、地下成对性状对不同或同一环境因子的差别响应与适应机制是成对性状之间关系变异的基础。
SLA-SRL对MAP、Ra以及IN的差别性响应,决定羌塘高原降水梯度带上二者强烈的权衡关系;MAP、Ra以及C∶N对LNarea产生显著影响,而与RNlength没有显著相关性,导致羌塘高原降水梯度带二者的权衡关系。在低降雨量和低养分环境中,SLA和叶片大小不断下降,而SRL保持不变或不断上升。例如,Holdaway等[50]沿土壤年代序列的研究发现,最高的SLA出现在土壤养分高的地点,而最高的SRL出现在土壤养分贫乏的区域;Liu等[29]沿降水梯度带研究发现SRL基本不变,而SLA变化显著。干旱胁迫对叶片性状具有很强的选择作用,而根系在水获取方面具有多方向策略。高辐射区域通常更干旱,MAP与Ra呈极显著负相关[18]。在更干旱、辐射更强的区域,植物叶片具有更低的SLA和更高的LNarea[69],从而提高水分利用效率[70]。LNmass-RNmass对MAP、Ra以及土壤C∶N具有相对一致的响应,羌塘高原降水梯度带二者呈极显著正相关,然而,LNmass与PET呈较显著负相关,RNmass与MAT呈显著负相关,对环境因子的差别响应导致羌塘高原降水梯度带二者具有显著的异速生长关系。PET与植物蒸散过程相关,在干旱环境中,植物通过降低叶片气孔导度减少水分散失[71-72],而为了补偿气孔数量及导度降低对光合效率的影响,干旱端具有更高的LNmass[55-57],维持更低水分和CO2导度下特定的光合速率[23];同时,干旱的条件下,植物通过增加叶片内部非光合器官或组织N的投入,提高细胞内部的渗透压,增强对体内水分的保护作用[73-74]。MAT直接影响高原冻土分布以及土壤冻融作用,而土壤冻融作用对根系限制较大,而对叶片影响较小。在低温环境下,植物地下部分趋向投入更多的N,增强根系的抵抗能力[48],同时,储存在根系里的养分也为第二年植物生长提供保障[75]。
地上、地下性状之间相关性不一致,说明植物地上、地下形态组分不同的环境压力[65]。不同环境条件下植物采取不同的资源利用策略,而不是对叶片和根系协同性状的一致性选择。根系属于结构器官,叶片属于代谢器官,二者养分含量受不同机制控制[17]。地上和地下性状可能存在根本不同的环境约束和选择压力[48]。干旱或极湿润端地上、地下成对性状具有更强的权衡策略,而中间区域高寒草甸的养分和水分限制不是很强烈,地上、地下性状表现出更多协同进化关系。羌塘高原降水梯度带成对性状的变异是干旱环境中物种的水分保持策略以及养分贫瘠环境中养分利用策略,涉及水分利用效率和N利用效率的权衡。
4 结论
羌塘高原环境梯度带为研究草地生态系统植物性状变异及其对高寒环境变化的适应策略提供了理想实验平台。在羌塘高原降水梯度带上,区域气候和土壤环境因子导致的叶片性状变异大于根系性状的变异,而根系性状的变异在物种间更大。在从西往东的降水量增加梯度带上,除了比叶面积呈现略微增加的趋势外,叶片和根系的养分含量都呈降低趋势,这种趋势在优势种物种水平更为明显。羌塘高原植物叶片和根系成对性状存在着权衡关系,在干旱的高寒草原、荒漠草原和极湿润的高寒草甸表现为更为显著的权衡关系,而在降水量的中间区的高寒草甸则表现为协同关系。地上、地下成对性状的变异特征及权衡关系体现了高寒植物对降水梯度带上环境因子变化的适应策略。干旱端植物通过增加比根长和叶片、根系养分含量来提高水分和养分的吸收能力,同时通过高叶片氮含量来维持光合碳获取能力,保障了根系生长的物质来源,表现出地上和地下同时投资的生态策略。干旱端植物保持较高的养分含量是抵御和适应严酷的寒、旱和贫瘠的环境胁迫的重要适应策略。而在湿润端植物则采取增加比叶面积,维持地上光合生产力的生态策略。本研究为理解高寒植物在环境梯度带的资源获取,以及优势种的广泛分布和生态适应策略提供了科学解释。
