敲减STAT3基因表达对胰腺祖细胞增殖的影响*
2020-03-03李旭艳翟文君马明君陈振宝孙玉林李永芹
李旭艳, 翟文君, 马明君, 陈振宝, 孙玉林, 赵 娟, 李永芹
(1岭南师范学院生命科学与技术学院, 广东 湛江 524000; 2东北林业大学生命学院, 黑龙江 哈尔滨 150040)
随着人们生活方式的改变和生活环境的变化,糖尿病和胰腺炎等胰腺相关疾病的发病率呈逐年上升趋势,严重危害人类健康和生活质量。糖尿病患者表现为胰岛β细胞受损或胰岛素抵抗,胰腺炎患者的胰腺腺泡细胞凋亡或坏死。补充受损或功能异常的胰岛β细胞或腺泡细胞将是治疗这类疾病的根本途径。终末分化胰岛细胞和胰腺腺泡细胞均起源于胚胎发育早期胰腺干细胞,研究发现,成体胰腺含有1%的胰腺干/祖细胞,激活成体胰腺干/祖细胞增殖分化,获得终末分化胰腺内、外分泌细胞,是胰腺疾病治疗的新希望。
信号转导及转录激活因子3(signal transducer and activator of transcription 3,STAT3)是重要的信号转导因子,参与干扰素、白细胞介素和生长因子等多种细胞因子的信号转导过程,是白细胞介素6/Janus 激酶(interleukin-6/Janus kinase,IL-6/JAK)和表皮生长因子受体(epidermal growth factor receptor,EGFR)等酪氨酸激酶信号通路效应分子,STAT3被上游的蛋白激酶磷酸化,形成二聚体,进入细胞核内调控促增殖、抗凋亡、免疫反应等基因表达,调控细胞生长、分化、迁移、凋亡、自噬、免疫以及代谢等重要生命活动。正常生理条件下STAT3低表达或短暂性激活,而在胰腺疾病发生发展过程中STAT3被活化并持续激活。被激活的STAT3促进胰腺癌细胞异常增殖、转移、生成新生血管[1],炎症在胰腺癌的进展过程中扮演重要角色,能显著加速胰腺上皮肿瘤癌变,STAT3作为一种急性期反应因子,参与调控胰腺炎和胰腺纤维化进程[2],另外,STAT3参与糖尿病肾病、糖尿病心肌病和糖尿病视网膜病变的发生和发展的过程[3-5]。阻断STAT3信号通路,能改善胰腺癌、胰腺炎和糖尿病并发症症状,STAT3信号通路有可能成为治疗胰腺疾病的药物靶点。研究发现,STAT3对肿瘤干细胞、胚胎干细胞和成体干细胞的增殖、自我更新和重编程有重要调控作用。STAT3促进牙髓干细胞增殖、抑制分化[6],促进甲状腺癌干细胞细胞集落形成[7],维持慢性髓细胞性白血病干细胞的特性[8],STAT3是否调控胰腺干/祖细胞的功能尚未报道。本研究以小鼠胰腺祖细胞为研究对象,沉默STAT3基因表达,检测STAT3基因对胰腺祖细胞增殖的影响,探究胰腺祖细胞增殖的机制,将为利用胰腺干/祖细胞开展疾病治疗提供实验依据。
材 料 和 方 法
1 细胞
小鼠成体胰腺祖细胞为东北林业大学发育生物学实验室提供[9]。
2 主要试剂
DMEM/F12培养液、胰蛋白酶、双抗和L-谷氨酰胺购自HyClone;B-27细胞培养添加剂和胎牛血清(fetal bovine serum,FBS)购自Gibco;细胞培养级别重组人胰岛素和表皮生长因子(epidermal growth factor,EGF)购自Becton, Dickinson and Company;细胞转染脂质体Lipofectamine RNAiMAX购自Invitrogen;总RNA提取试剂Tripure和实时荧光定量PCR试剂RT-qPCR SYRB Green购自Roche;脱氧核糖核酸酶I(deoxyribonuclease I,DNaseI)和核糖核酸酶抑制剂(ribonuclease inhibitor,RI)购自TaKaRa;反转录试剂盒、抗β-肌动蛋白(β-actin)抗体(HC201-02)和辣根过氧化物酶标记的II抗(HS201-01)购自北京全式金生物技术有限公司;Cell Counting Kit-8(CCK8)试剂盒购自Dojindo Labratories;细胞周期检测试剂盒、BCA蛋白浓度测定试剂盒和RIPA蛋白裂解液购自碧云天生物技术研究所;抗STAT3抗体(cst.9139)和抗细胞周期蛋白D2(cyclin D2,CCND2)抗体(cst. 3741)购自Cell Signaling Technology;β-巯基乙醇、过硫酸铵、丙烯酰胺、甲基双丙烯酰胺和四甲基乙二胺购自Sigma;吐温20、甲醇等常规生化试剂购自天津天大化学试剂厂。靶向STAT3的小干扰RNA[STAT3small interfering RNA (siRNA),siSTAT3][10]和对照siRNA(negative control,NC)由吉玛基因股份有限公司合成,序列见表1。qPCR引物由金唯智基因公司合成,序列见表2。

表1 siRNAs序列
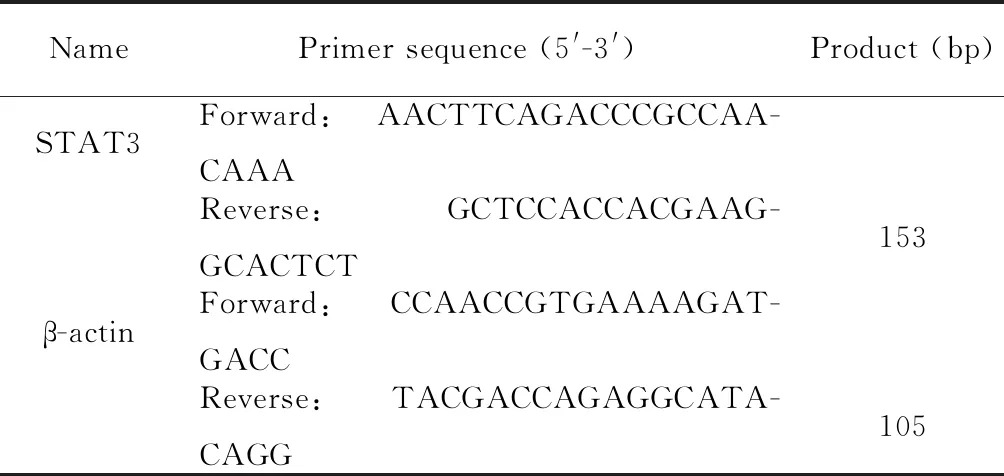
表2 qPCR引物序列
3 主要方法
3.1细胞培养 传代胰腺祖细胞用10%贴壁培养液过夜培养,次日早更换2%生长培养液,置于37 ℃、5.0% CO2饱和湿度的培养箱中培养,每3 d更换生长培养液。贴壁培养液成分:含10% FBS、1%双抗和1% L-谷氨酰胺的DMEM/F12培养液。生长培养液成分:含2% FBS、1%双抗、1% L-谷氨酰胺、2% B-27、50 μmol/L β-巯基乙醇、10 mg/L重组人胰岛素和20 μg/L EGF的DMEM/F12培养液。
3.2细胞转染 取培养8代的胰腺祖细胞,96 孔和12 孔细胞培养板每孔分别接种3×103和1×105个细胞,生长培养液培养24 h后,细胞40%汇合后,每孔转染50 nmol/L siSTAT3或NC,细胞转染参照Lipofectamine RNAiMAX说明书操作。对照组和实验组均设3个复孔。同时转染阳性对照siRNA,qPCR检测基因沉默效果, 间接判断转染效率。
3.3qPCR检测各基因mRNA的表达 细胞转染48 h,提取细胞总RNA,利用随机引物反转录,获得cDNA,qPCR检测基因mRNA表达,所得数据通过2-ΔΔCt方法计算,β-actin作为内参照,设置3个重复。
3.4Western blot检测蛋白的表达 细胞转染72 h,PBS洗涤,RIPA裂解细胞,收集裂解液,10 000×g离心10 min,取上清,测定蛋白浓度,分装样品冻存。样品进行SDS-PAGE,转膜,37 ℃封闭1 h,抗β-actin、STAT3和CCND2 抗体4 ℃孵育过夜,TBST洗膜3次,II抗室温孵育1 h,Tanon 5200发光成像系统采集图像,β-actin作为内参照。
3.5活细胞计数检测细胞增殖 24孔板接种胰腺祖细胞,生长至40%汇合,转染siSTAT3或NC 72 h,胰酶消化为单细胞,收集到离心管定容到1 mL,稀释后,在光学显微镜下用血细胞计数板计数。对照组和实验组均设3个复孔。
3.6CCK8法检测细胞活力 96 孔板接种胰腺祖细胞,生长至40%汇合,转染siSTAT3或NC 72 h,弃去培养液,PBS洗涤细胞,补加110 μL含有10% CCK8的DMEM/F12培养液,置于37 ℃培养箱中培养2 h,通过酶标仪读取各孔吸光度(A)值。对照组和实验组均设3个复孔。
3.7流式细胞术检测细胞周期分布 细胞转染72 h,收集各组细胞1×106个,PBS洗涤2次,1 100×g离心5 min,70%冷乙醇4 ℃固定2 h,PBS洗2 次,30 mg/L RNAse A 37 ℃孵育30 min,50 mg/L PI避光孵育30 min,冷PBS洗2次,600 μL PBS重悬细胞,Accuri C6流式细胞仪检测,随机收集30 000个细胞,采集信号,采用ModFit 4.1流式数据分析软件进行数据分析。
4 统计学处理
用GraphPad Prism 7软件进行统计学处理,数据采用平均数±标准误(mean±SEM)表示,两组间比较采用独立样本t检验,以P<0.05为差异有统计学意义。
结 果
1 敲减STAT3基因表达的效果
50 nmol/L siSTAT3或NC转染胰腺祖细胞48 h和72 h后,分别提取RNA和蛋白,通过qPCR和Western blot检测siRNA对STAT3基因的沉默效果。与对照组相比,实验组胰腺祖细胞STAT3的mRNA和蛋白表达水平明显下调(P<0.05),mRNA的表达水平降低了(49.75±0.03)%,蛋白的表达水平降低了(78.62±0.08)%,见图1。

Figure 1. The relative expression ofSTAT3gene in the pancrea-tic progenitor cells transfected withSTAT3siRNA. A: the mRNA expression of STAT3 was detected by qPCR after transfection for 48 h; B: the protein expression of STAT3 was determined by Western blot after transfection for 72 h. Mean±SEM.n=3.**P<0.01vsNC group.
图1 转染STAT3siRNA后胰腺祖细胞STAT3基因表达水平的变化
2 敲减STAT3基因表达对胰腺祖细胞增殖和活力的抑制作用
细胞转染后,倒置显微镜下观察,转染72 h后,采集细胞图像结果显示,转染STAT3siRNA的胰腺祖细胞孔板中细胞密度降低,见图2A;活细胞计数结果显示,敲减STAT3基因的表达使细胞数量减少了(45.62±0.03)%(P<0.01),见图2B。CCK8法检测结果显示敲减STAT3基因的表达使胰腺祖细胞活力减少了(43.68±0.05)%(P<0.05),见图2C。
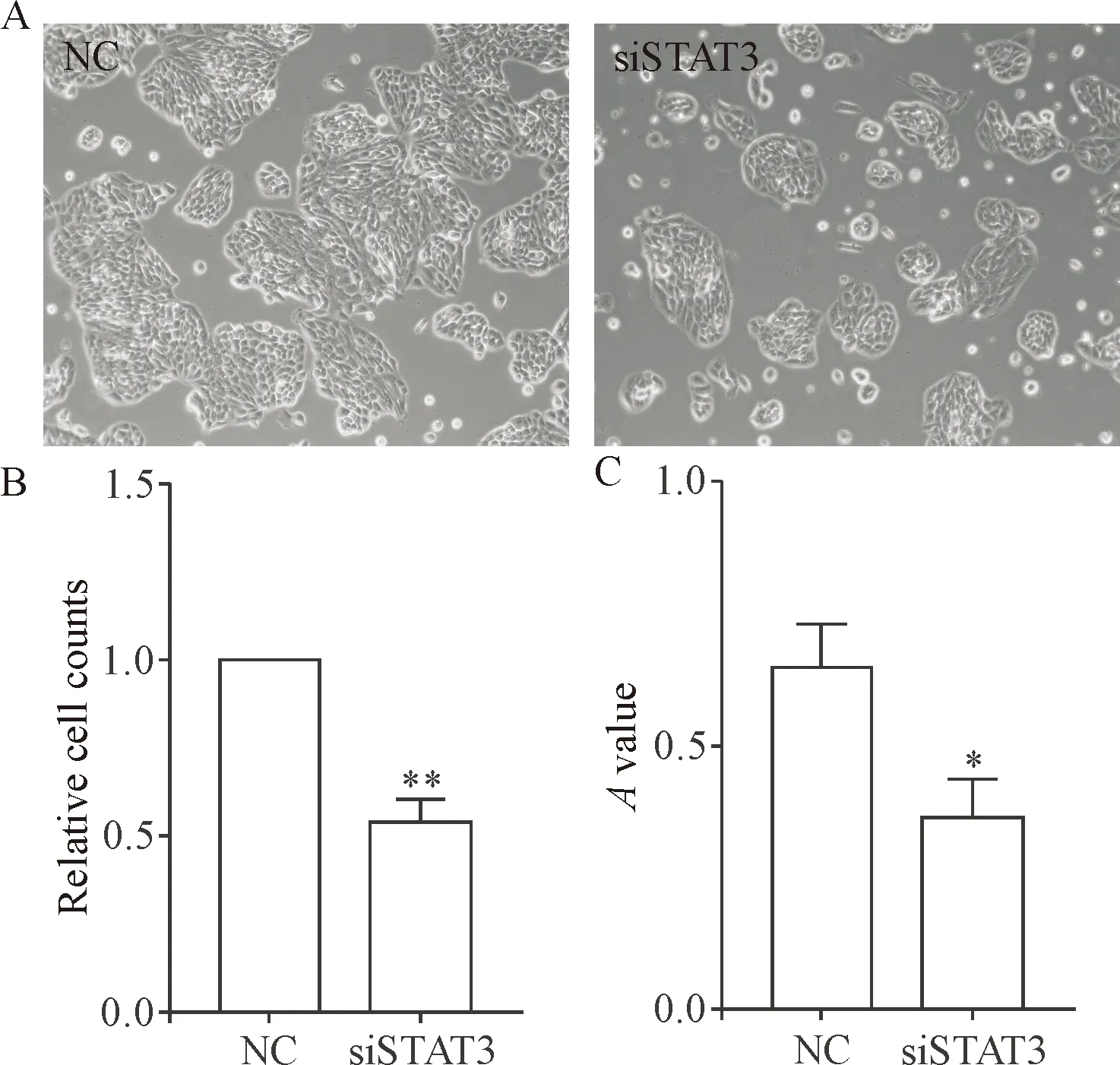
Figure 2. The effects ofSTAT3silencing on the proliferation and viability of pancreatic progenitor cells. A: the images of cells under the inverted optical microscope after transfection for 72 h (×100); B: the effect ofSTAT3silencing on the proliferation of pancreatic progenitor cells was detected by cell counting after transfection for 72 h; C: the effect ofSTAT3silencing on the viability of pancreatic progenitor cells was measured by CCK8 assay after transfection for 72 h. Mean±SEM.n=3.*P<0.05,**P<0.01vsNC group.
图2 沉默STAT3对胰腺祖细胞增殖和活力的影响
3 敲减STAT3的表达对胰腺祖细胞细胞周期分布的影响
细胞转染72 h,经流式细胞术检测细胞周期分布。与对照组相比,转染siSTAT3干扰片段的胰腺祖细胞其周期阻滞在G0/G1期,见图3。
4 敲减STAT3的表达对CCND2蛋白表达的影响
细胞转染72 h,Western blot 检测结果可见转染siSTAT3的胰腺祖细胞中CCND2蛋白表达水平明显降低(P<0.05),见图4。
讨 论
糖尿病发病率呈逐年上升趋势,利用胚胎、多能性、成体干细胞诱导生成分泌胰岛素的细胞是糖尿病干细胞治疗的主要研究方向,激活内源胰腺干/祖细胞生成功能细胞,将是一种更为有效和安全的治疗方案,然而成体胰腺干/祖细胞数量少且常处于静息状态,因此,探究胰腺干/祖细胞的增殖机制具有重要的科学意义。研究表明,STAT3 在不同类型的细胞和组织中广泛表达,参与细胞生长和凋亡等功能的调控,本研究利用RNAi技术成功敲减胰腺祖细胞STAT3基因表达,细胞计数和CCK8检测结果表明,敲减STAT3抑制胰腺祖细胞增殖和细胞活力。
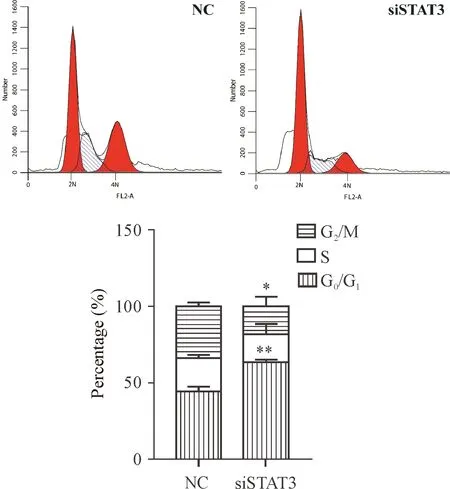
Figure 3. The effects ofSTAT3silencing on the cell cycle distribution of pancreatic progenitor cells were analyzed by flow cytometry after transfection for 72 h. Mean±SEM.n=3.*P<0.05,**P<0.01vsNC group.
图3 流式细胞术检测沉默STAT3对胰腺祖细胞细胞周期分布的影响

Figure 4. The effects ofSTAT3silencing on the protein expression of CCND2 in the pancreatic progenitor cells after transfection for 72 h. Mean±SEM.n=3.*P<0.05vsNC group.
图4 沉默STAT3对胰腺祖细胞CCND2表达的影响
STAT3 是转录激活因子家族成员之一,在多种信号分子刺激下,STAT3被磷酸化激活,结合于靶基因DNA调控区,调控功能基因表达。STAT3活化后可通过激活其靶基因MYC原癌基因、细胞周期蛋白D1(cyclin D1,CCND1)、CCND2、存活蛋白(survivin)、基质金属蛋白酶2(matrix metalloproteinase-2,MMP2)、p21、p27等,调控细胞增殖、细胞周期进程、迁移、侵袭和凋亡[11-12]。通过慢病毒介导的短发夹RNA(short hairpin RNA,shRNA)沉默STAT3基因表达,可降低survivin基因表达,抑制成牙骨质细胞分化,诱导细胞凋亡[13]。人工合成STAT3蛋白抗体SBT-100特异性抑制磷酸化STAT3功能,可抑制乳腺癌细胞增殖[14];siRNA沉默STAT3基因表达可降低survivin和CCND1表达,抑制白细胞介素17(interleukin-17,IL-17)对人角质形成细胞增殖的诱导作用[15]。研究表明,STAT3还通过调控下游基因表达影响干细胞增殖、自我更新和分化[16]。已报道,减弱JAK/STAT3信号活性抑制骨髓基质细胞增殖和成骨分化[17]。激活STAT3基因,促进小鼠胚胎干细胞多潜能性调控因子MYC和Bcl-3表达,有利于维持细胞自我更新特性[18-19]。STAT3促进CD24、CD34、CD38、CD44、CD90和CD133等肿瘤干细胞标志性基因表达[20],STAT3与CD44形成二聚物并乙酰化,入核后结合CCND2、MYC和TWIST1基因启动子,激活细胞周期调控基因表达[21-22]。本研究显示,在胰腺祖细胞中敲减STAT3基因可降低CCND2蛋白表达,使细胞周期进程阻滞在G0/G1期,抑制胰腺祖细胞细胞增殖。这表明,STAT3基因通过调控细胞周期蛋白CCND2的表达调控胰腺祖细胞增殖。
STAT3磷酸化能被多种途径激活。EGF、血管内皮生长因子等与相应受体结合,使受体磷酸化,利用其内源性酪氨酸激酶活性直接磷酸化STAT3;IL-6等细胞因子与细胞膜上相应受体结合,募集JAK使受体的酪氨酸残基磷酸化,募集STAT3并使其磷酸化。 研究表明,EGF是小鼠和人胚胎胰腺祖细胞、小鼠成体胰腺祖细胞维持体外增殖、自我更新和诱导分化的必需因子,用缺乏EGF的培养液培养胰腺祖细胞,使细胞增殖速度减慢[23-24]。EGF与其受体EGFR结合,使受体磷酸化,激活促进细胞增殖和细胞存活的信号通路,如JAK/STAT3、丝裂原活化蛋白激酶/细胞外信号调节激酶(mitogen-activated protein kinase/extracellular signal-regulated kinase,MAPK/ERK)和磷酯酰肌醇3-激酶/蛋白激酶B(phosphatidylinositol 3-kinase/protein kinase B,PI3K/AKT)信号通路。我们推测,敲减STAT3基因表达可能阻断了上游EGFR/JAK/STAT3信号通路的传导作用,从而抑制细胞增殖。
STAT3已成为转录因子、微小RNA(microRNA,miRNA)、长链非编码RNA(long non-coding RNA,lncRNA)、小分子抑制剂和药物调控细胞功能的靶点,如抗肿瘤药CDK5抑制剂Roscovitine对大鼠血管平滑肌细胞增殖的抑制作用,以及lncARSR(lncRNA activated in renal cell carcinoma with sunitinib resis-tance)对肝癌干细胞增殖的促进作用是通过调控STAT3途径实现的[25-26]。STAT3信号通路在胰腺癌、胰腺炎和糖尿病并发症中发挥重要功能,STAT3信号通路可能成为胰腺疾病治疗的靶点。本研究揭示了STAT3调控胰腺祖细胞增殖的机制,为利用胰腺干/祖细胞开展疾病治疗提供了实验依据。
