我国朱槿曲叶病毒病及其传播介体烟粉虱分布调查
2020-02-22陈婷汤亚飞何自福吕利华齐国君
陈婷 汤亚飞 何自福 吕利华 齐国君


摘要:【目的】明確我国朱槿植株上木尔坦棉花曲叶病毒(Cotton leaf curl Multan virus, CLCuMuV)引起的朱槿曲叶病发生情况及烟粉虱隐种种群组成,为朱槿曲叶病的早期监测及基于控制传播介体的病毒防控提供科学依据。【方法】2017年11月—2018年8月,从我国广东、广西、海南、云南、福建、新疆、内蒙、山东、浙江和四川等10省(区)32市(县)的城市绿化区域(公园、公路旁绿化区域)及花卉市场采集朱槿曲叶病疑似病株及健康朱槿植株叶片,运用PCR进行CLCuMuV检测,并利用mtCOⅠ基因序列对采自田间朱槿植株上的单头烟粉虱进行隐种鉴定及UPGMA聚类分析。【结果】我国广东、广西、海南、云南和福建5省(区)绿化朱槿被CLCuMuV侵染发病的比率较高,平均为44.43%~81.95%,高于新疆(4.39%)和内蒙古(0.63%)的发病率,而山东、浙江和四川的朱槿植株暂未发现CLCuMuV感染。在大部分采样点,烟粉虱种群密度较低,除内蒙古的平均虫株率为15.00%外,其他省(区)的平均虫株率均低于5.00%。对采集的烟粉虱隐种鉴定结果表明,烟粉虱种群包括MEAM1、MED、AsiaⅡ7、AsiaⅡ1和AsiaⅠ等5个隐种,其中AsiaⅡ7和MEAM1为优势种群,所占比例分别为44.7%和32.5%。【结论】目前我国南方地区朱槿曲叶病发病率较高,其传毒介体烟粉虱AsiaⅡ7和MEAM1为优势种群。随着朱槿盆栽花卉贸易及苗木跨区调运,CLCuMuV有向内地逐渐扩散的趋势,应采取相应措施提早防范木尔坦棉花曲叶病毒在我国棉区发生流行。
关键词: 朱槿;木尔坦棉花曲叶病毒;烟粉虱;发生分布
中图分类号: S436.81 文献标志码: A 文章编号:2095-1191(2020)11-2697-09
Distribution investigation of Cotton leaf curl Multan virus on Hibiscus rosa-sinensis and Bemisia tabaci Gennadius in China
CHEN Ting, TANG Ya-fei, HE Zi-fu, LYU Li-hua*, QI Guo-jun*
(Plant Protection Research Institute, Guangdong Academy of Agricultural Sciences/Guangdong Provincial Key
Laboratory of High Technology for Plant Protection , Guangzhou 510640, China)
Abstract:【Objective】The objective of this study was to identify the occurrence of Cotton leaf curl Multan virus (CLCuMuV) and the Bemisia tabaci Gennadius on Hibiscus rosa-sinensis plants in China, provide early warning and monitoring for the cotton leaf curl disease caused by this virus, and provide a basis for preventing the outbreak of the vector and diseases.【Method】Using PCR method, the CLCuMuV detection on suspected diseased plants and healthy plants of hibiscus collected from urban green areas(park, roadside green areas) and flower markets in 32 cities/counties of 10 provinces/regions(Guangdong,Guangxi,Hainan,Yunnan,Fujian,Xinjiang,Inner Mongolia,Shandong,Zhejiang and Sichuan) of China in 2017-2018 were detected. mtCOⅠgene sequence was used for identification of whitefly collected from the hibiscus, and the UPGMA phylogenetic tree was constructed by using MEGA 5.0. 【Result】 The hibiscus infected CLCuMuV were serious in five provinces(regions):Guangdong, Guangxi, Hainan, Yunnan and Fujian , the average incidence rate was 44.43%-81.95%, which was higher than Xinjiang(4.39%) and Inner Mongolia(0.63%). Hibiscus had not been found infected CLCuMuV in Shandong, Zhejiang and Sichuan. In most of the sampling sites, the population density of whitefly was low. The average whitefly in Inner Mongolia was 15.0%, and other provinces(regions) was lower than 5.00%. The collected whiteflies were identified based on mtCOⅠgene sequence, the results showed that there were five cryptic species, including MEAM1, MED, AsiaⅡ7 , AsiaⅡ1 and AsiaⅠ. AsiaⅡ7 and MEAM1 were the dominant species, the proportions were 44.7% and 32.5% respectively. 【Conclusion】CLCuMuV has been occurring seriously on hibiscus in southern China at present, the whitefly AsiaⅡ7 and MEAM1 are the dominant species on hibiscus. With the cross-regional transport of the CLCuMuV-infected hibiscus plants, CLCuMuV has a gradual spread and epidemic trend in China.
Key words: Hibiscus rosa-sinensis; Cotton leaf curl Multan virus; Bemisia tabaci Gennadius ; occurrence and distribution
Foundation item: National Natural Science Foundation of China(31871937, 32001973); Guangdong Natural Science Foundation(2020A1515011098); Guangzhou Science and Technology Project(201904010173)
0 引言
【研究意义】烟粉虱(Bemisia tabaci Gennadius)传双生病毒(Whitefly-transmitted geminiviruses,WTGV)病是世界范围内广泛发生流行的主要病害之一,对多种重要经济作物生产造成严重损害,具传毒介体烟粉虱在双生病毒的侵染传播过程中发挥着至关重要的作用。棉花曲叶病是棉花上的一种毁灭性病害,已给南亚地区棉花生产造成巨大损失(Maharshi et al.,2017),木尔坦棉花曲叶病毒(Cotton leaf curl Multan virus,CLCuMuV)是引起该病害的主要病原之一(Briddon and Markham,2000;Ahmed et al.,2011),已入侵我国并在多种锦葵科植物上有报道(董迪等,2010;Cai et al.,2010;汤亚飞等,2013,2015a,2015b)。CLCuMuV可侵染中间寄主,并通过介体烟粉虱传播到棉花作物上(Maharshi et al.,2017),一旦携毒烟粉虱扩散蔓延至棉花主产区,其暴发流行将对我国的棉花生产造成毁灭性危害。在我国,朱槿是CLCuMuV主要寄主植物(何自福等,2012;陈婷等,2016a),因此,探明我国朱槿曲叶病的发生分布情况及烟粉虱隐种种群结构及区域分布,分析CLCuMuV流行傳播及发生风险,对该病毒早期预警及基于控制传播介体的防控具有重要意义。【前人研究进展】国际上已鉴定出引起棉花曲叶病的双生病毒有10种,其中CLCuMuV及其伴随的卫星分子是20世纪90年代在南亚地区引起棉花曲叶病的主要病原微生物(Briddon et al.,2003;Mansoor et al.,2003;Sattar et al.,2013)。虽然朱槿曲叶病在印度北部最早报道于1950年,但直到2005年Rajeshwari等才通过系统发育分析明确了印度朱槿上的病毒分离物与CLCuMuV的相似性在95%~97%。在我国,2006年在广东省的朱槿上最先发现CLCuMuV(毛明杰等,2008),现已在广西、海南、福建、云南和江苏等地发现该病毒侵染为害黄秋葵(董迪等,2010)、棉花(Cai et al.,2010;汤亚飞等,2015a)、垂花悬铃花(汤亚飞等,2013)、朱槿(章松柏等,2013;张晖等,2015)和红麻(汤亚飞等,2015b)等锦葵科植物。Du等(2015)分析了我国与南亚CLCuMuV的系统发育关系,推断从南亚输入的朱槿为入侵我国的CLCuMuV来源。烟粉虱是CLCuMuV的唯一传播昆虫介体(刘银泉和刘树生,2012)。在南亚棉花曲叶病暴发区域,烟粉虱隐种包括MEAM1及AsiaⅡ1、AsiaⅠ、AsiaⅡ5、AsiaⅡ7和AsiaⅡ8(Dinsdale et al.,2010;Ashfaq et al.,2014)。在我国华南地区,朱槿上的烟粉虱隐种主要为MEAM1和AsiaⅡ7隐种(陈婷等,2016a)。室内传毒试验证明入侵隐种MEAM1和MED不传播CLCuMuV,本地隐种AsiaⅡ1和AsiaⅡ7可传播CLCuMuV致使棉花染病(Pan et al.,2018;Chen et al.,2019)。【本研究切入点】朱槿作为重要的园林绿化植物及家庭盆栽花,种植十分广泛,目前华南地区朱槿已普遍感染CLCuMuV(陈婷等,2016a),并已形成长期而稳定的毒源库(何自福等,2012),随着携毒朱槿苗木跨区调运和传毒介体烟粉虱扩散,CLCuMuV向我国棉区传播扩散的潜在风险急剧增大,但目前尚无针对我国朱槿曲叶病发病率、烟粉虱隐种组成及发生分布情况的调查研究报道。【拟解决的关键问题】全面普查我国城市绿化区和花卉市场朱槿植株上CLCuMuV侵染情况及烟粉虱隐种的发生与分布,明确该病毒和传毒烟粉虱隐种在我国的发生为害情况,以期为朱槿曲叶病的早期监测及基于控制传播介体的病毒防控提供科学依据。
1 材料与方法
1. 1 试验材料
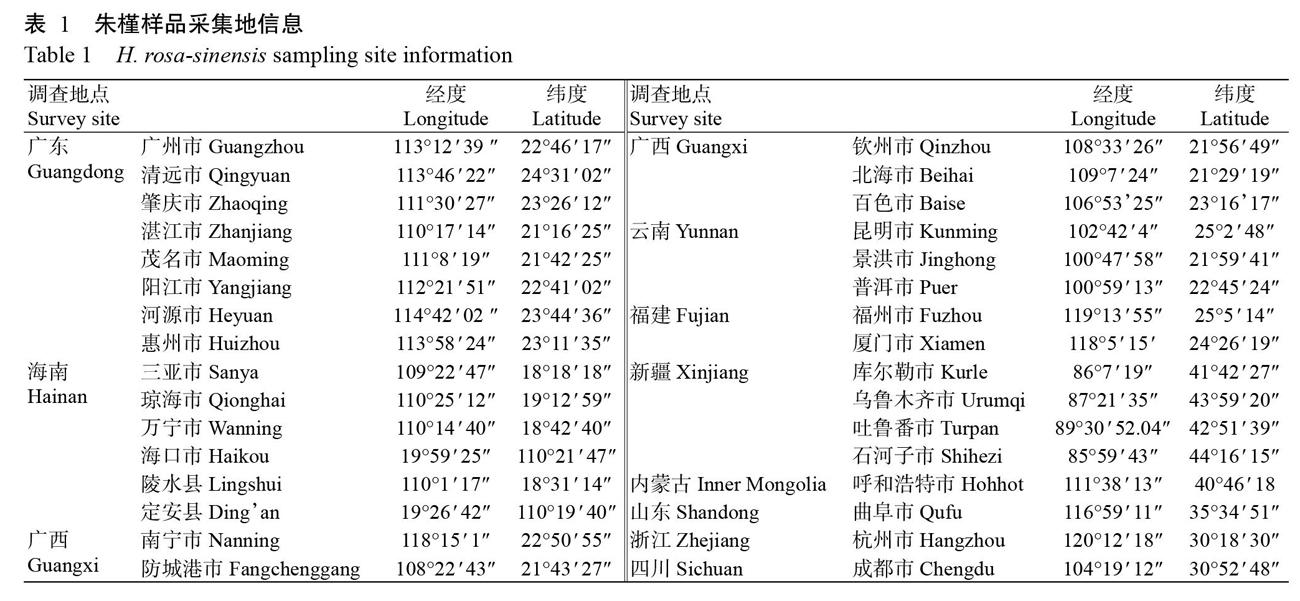
1. 1. 1 样品来源 2017年11月—2018年8月,从我国10省(区)32市(县)的城市绿化区域(公园、公路旁绿化区域)及花卉市场采集朱槿叶片,用于CLCuMuV检测。田间朱槿植株上的烟粉虱样品共计351头,采集自7省(区)18市(县)。采集地信息见表1。
1. 1. 2 主要试剂及仪器 植物细胞组织基因组试剂盒EF111-12(北京全式金生物技术有限公司);PCR Mixer(日本TaKaRa公司);其他试剂均为国产分析纯。引物由生工生物工程(上海)股份有限公司合成;由上海英骏生物技术有限公司完成测序。主要仪器设备:Mastercycler梯度PCR仪(德国Eppendorf公司)、Gel Doc XR凝胶成像系统(美国Bio-Rad公司)和DYY-12电泳仪(北京六一仪器厂)。
1. 2 试验方法
1. 2. 1 朱槿曲叶病发病率调查及叶片取样 在每个地区选取3~5个调查点,记录朱槿的发病株及健康株,计算发病率。每个调查点公园绿化区调查面积100 m2,道路绿化区调查长度约100 m。调查标准:朱槿全株叶片黄化、叶片向上卷曲、叶脉肿大明显,并且产生叶耳、开花少或不开花为病株,全株叶片无上述症状即为健康植株(汤亚飞等,2015b)。在每个调查点分别采集7株显症朱槿叶片和未显症叶片,放入塑料密封袋中,标记采集地点及时间。所有采集的样品带回实验室,放入-80 ℃超低温冰箱保存备用。
1. 2. 2 朱槿上烟粉虱成虫采集 朱槿曲叶病调查及取样时用吸虫器采集烟粉虱成虫。将采集的标本放入装有95%乙醇的2 mL离心管中,带回实验室,置于-20 ℃冰箱中保存备用。
1. 2. 3 朱槿叶片CLCuMuV检测 从-80 ℃超低温冰箱取出样本,采用植物细胞组织基因组试剂盒EF111-12提取朱槿植株叶片总DNA,具体操作按试剂盒说明进行。采用CLCuMuV特异引物CL-F、CL-R和β卫星分子特异引物beta-F、beta-R(表2)分别进行PCR检测。PCR反应体系及扩增程序参见汤亚飞等(2015a)的方法。
1. 2. 4 烟粉虱隐种鉴定 将保存的烟粉虱样本取出,参考Frohlich等(1999)的方法对单头烟粉虱DNA提取扩增,进行隐种鉴定。吸取30~40 μL裂解液[裂解液浓度:5 mmol/L Tris(pH 8.0),0.5 mmol/L EDTA,0.5% NP-40,1 mg/mL蛋白酶K;裂解液体积:5 mmol/L Tris (pH 8.0) 5 μL,0.5 mmol/L EDTA 1 μL,0.5% NP-40 5 μL,ddH2O 989 μL],单头烟粉虱置于其中1滴裂解液中。用0.2 mL PCR管底部将虫体充分研磨匀浆,将匀浆液吸入1.5 mL离心管内。再吸取另外1滴裂解液清洗Parafilm膜,将清洗液与匀浆液混合后置于65 ℃水浴15~25 min,随后95 ℃ 10 min,使蛋白酶K失活,短暂低速离心后即可用于PCR。利用烟粉虱mtCOⅠ基因通用引物C1-J-2195和TL2N-3014(表2)对单头烟粉虱DNA进行PCR检测,PCR反应体系及扩增程序参见陈婷等(2016b)的方法。将获得的烟粉虱mtCOⅠ基因序列,利用DNASTAR进行序列拼接,并通过序列同源性比对分析对烟粉虱隐种进行鉴定。
1. 2. 5 烟粉虱系统发育分析 将测序所得烟粉虱mtCOⅠ基因序列在NCBI中进行BLAST分析。利用MEGA 5.0进行UPGMA聚类分析,构建系统发育进化树。
1. 3 统计分析
试验数据使用Excel 2010进行统计分析。
2 结果与分析
2. 1 朱槿曲叶病发病率调查结果
对我国10省(区)32市(县)朱槿叶片携毒检测结果(表3)表明,广东、广西、海南、福建和云南5省(区)的朱槿感染CLCuMuV情况较重,朱槿曲叶病发病率较高,平均发病率分别为81.95%、66.26%、58.75%、51.05%和44.43%;新疆花卉市场及花卉生產大棚的朱槿仅有少量感染曲叶病,平均发病率为4.39%;内蒙古花卉市场的朱槿有极少量感染曲叶病,发病率仅为0.63%。此外,山东城市绿化区及浙江、四川花卉市场的朱槿植株未发现曲叶病。可见,不同地区朱槿感染CLCuMuV的程度明显不同,其中广东、广西、海南、福建和云南5省(区)的朱槿发病率高于新疆和内蒙古地区,而山东、浙江和四川暂未发现朱槿感染CLCuMuV。
2. 2 朱槿上烟粉虱虫株率调查结果
朱槿植株上的烟粉虱发生调查结果(表3)表明,10省(区)32市(县)中有7省(区)18市(县)的朱槿上有烟粉虱发生,但烟粉虱虫株率相对较低。广东、海南、广西、云南和福建5省(区)园林绿化朱槿上烟粉虱平均虫株率分别为1.84%、0.98%、1.33%、1.90%和4.05%;新疆花卉市场及花卉生产大棚的烟粉虱平均虫株率为2.25%;内蒙古呼和浩特花卉市场的朱槿上烟粉虱平均虫株率为15.0%。山东城市绿化区及浙江、四川花卉市场的朱槿植株上未发现烟粉虱。可见,56.25%的采样地点有烟粉虱发生,但烟粉虱种群密度较低,平均虫株率低于5.00%,且不同地区的烟粉虱发生情况略有差异。
2. 3 朱槿上烟粉虱隐种种类及比例
利用mtCOⅠ基因片段序列对烟粉虱隐种进行鉴定,结果(表4)表明,7省(区)18市(县)共发现5个烟粉虱隐种,主要包括入侵隐种MEAM1、MED和本地隐种AsiaⅡ7、AsiaⅡ1和AsiaⅠ。在采集到的烟粉虱混合隐种种群中,本地隐种AsiaⅡ7为优势种,所占比例达44.7%,其次是入侵隐种MEAM1,所占比例为32.5%,再次为入侵隐种MED,所占比例为19.4%,AsiaⅠ和AsiaⅡ1本地隐种的数量较少,所占比例分别为2.9%和0.6%。从不同省(区)烟粉虱隐种组成情况看,广东主要是MEAM1和AsiaⅡ7;海南有4个隐种,包括AsiaⅠ、AsiaⅡ7、MEAM1和MED;云南和福建均只有AsiaⅡ7;广西为AsiaⅡ7和AsiaⅡ1;新疆主要为MED,还有少量的AsiaⅡ7和MEAM1;内蒙古主要为MED。
2. 4 朱槿上烟粉虱系统发育分析结果
为比较我国朱槿上烟粉虱与其他烟粉虱隐种间的关系,利用MEGA 5.0构建系统发育进化树。选取NCBI上已登录的烟粉虱部分隐种的mtCOⅠ基因序列作为参考序列,包括AsiaⅡ7(KM821541和DQ116650)、AsiaⅡ1(KR020527)、AsiaⅠ(KR 110156)、MEAM1(AJ550176)和MED(FN557454和KT235801),结合本研究获得的我国18个市(县)朱槿上24条不同的烟粉虱mtCOⅠ基因序列,并以温室白粉虱(AF418672)和非洲伯粉虱(AJ784260)为外群,进行系统发育进化树构建。结果(图1)显示,烟粉虱mtCOⅠ基因序列根据隐种分成7个组,本研究所得到的5个烟粉虱隐种基因序列均与已报道隐种各自聚为一组,全国各地不同地点的同一隐种均聚为一组。2个外群各自聚为一组。
3 讨论
3. 1 我国朱槿曲叶病调查
本研究对我国10省(区)朱槿上的CLCuMuV进行调查及检测,结合目前我国对CLCuMuV的相关报道发现,自2006年首次在广州市发现CLCuMuV侵染的朱槿表现出曲叶病症状以来,短短几年时间就在广东、广西、福建和海南等地普遍发现朱槿曲叶病。2007—2009年,CLCuMuV引起的朱槿曲叶病在广西南宁扩散蔓延加剧(林林等,2011);2008年,在南宁的田间发现CLCuMuV侵染的棉花植株(Cai et al.,2010)。2011—2012年,在福建厦门、福州和宁德等地的公路绿化带发现CLCuMuV引起的朱槿曲叶病株(章松柏等,2013)。2012年,在江苏南京的朱槿(张晖等,2015)和海南的红麻(汤亚飞等,2015b)上鉴定出CLCuMuV。在国外,早在1980年就有在印度北部朱槿上发生曲叶病的报道(Mali,1980),且在印度南部种植的101个朱槿品种中有75个品种极易感染CLCuMuV(Rajeshwari et al.,2005);2012年,在菲律宾也发现CLCuMuV侵染朱槿,且与我国广东的该病毒分离物GD01(KP762786)的相似度最高(She et al.,2017)。可见,CLCuMuV正在经历由点到面的发展、扩散。调查表明广东、广西、海南、云南和福建5省(区)的朱槿曲叶病发生率较高,个别城市的朱槿曲叶病发生率高达100.00%。广东、广西、海南、云南和福建地区处于朱槿的适生区域,气候环境利于朱槿生存,并且与前人预测的棉花曲叶病在我国的潜在适生区相吻合(齐国君等,2016)。通过走访各市(县)花卉市场的卖主和苗圃花农得知,朱槿苗木大多由广东或福建调运而来,随后经过枝条扦插繁殖而扩大种植。山东、浙江和四川暂未发现CLCuMuV,可能是因为调运的朱槿苗木中未携带CLCuMuV,而新疆和内蒙古花卉市场发现的朱槿曲叶病植株可能来自于从发病地区调运的带病植株。因此,应密切关注CLCuMuV的流行发生趋势,以防造成更严重的危害。
3. 2 我国朱槿上烟粉虱调查
烟粉虱是CLCuMuV的主要传播途径之一,监测植株上的传毒媒介是预防棉花曲叶病毒病发生的关键。传毒烟粉虱隐种在棉花曲叶病流行中具有不可忽视的作用。室内传毒已验证AsiaⅡ1和AsiaⅡ7烟粉虱可将棉花上的CLCuMuV传播至红麻、黄秋葵和棉花等锦葵科植物引起曲叶病(陈婷等,2016b;Pan et al.,2018;Chen et al.,2019)。巴基斯坦的棉花曲叶病发生地调查发现,曲叶病发病率高的地区烟粉虱主要是AsiaⅡ1隐种,而发病率低地区的烟粉虱主要是MEAM1隐种(Ahmed et al.,2011)。AsiaⅡ1隐种是引起棉花曲叶病在巴基斯坦田间流行的主要烟粉虱隐种。本研究对朱槿上烟粉虱的发生及隐种鉴定结果表明,我国朱槿上的烟粉虱共5个隐种,包括入侵隐种MEAM1、MED和本地隐种AsiaⅡ7、AsiaⅡ1、AsiaⅠ,其中,本地隐种AsiaⅡ7和入侵隐种MEAM1所占比例最高。因此,烟粉虱AsiaⅡ7隐种是我国CLCuMuV在田间扩散的途径之一。
煙粉虱AsiaⅡ7最早在广州市报道(邱宝利等,2006),MEAM1虽然是世界性的入侵隐种,对另一本地隐种ZHJ1(AsiaⅡ3)具有较强的取代能力(Liu et al.,2007),但MEAM1与AsiaⅡ7的竞争能力尚未知。但与蔬菜等作物相比,AsiaⅡ7在园林植物上适合度更高(林莉等,2010;Qiu et al.,2011),对棉花、黄秋葵等锦葵科寄主植物的趋性也较番茄、茄子等蔬菜作物更强(陈婷等,2019)。因此,即使入侵隐种MEAM1是我国南方地区的主要烟粉虱隐种(陈婷等,2016b),AsiaⅡ7隐种独特的生态位,可能是该隐种与MEAM1虽然同时存在于朱槿植株上,但AsiaⅡ7隐种是朱槿植株上的优势烟粉虱隐种所致。用UPGMA方法构建的系统发育进化树表明,我国各地的AsiaⅡ7烟粉虱基因序列聚为一支,究其原因是:对于地理距离较近的种群,可能因小气候环境较相似及自然扩散作用促进各种群间的基因交流;而如库尔勒等地理距离相对较远的地理种群也聚为一支的现象,很可能是这些烟粉虱种群随南方地区引进的花卉苗木调运至当地,温室大棚的环境适宜其生存,且因数量较少不易被察觉。
3. 3 朱槿曲叶病毒病与烟粉虱的相关性及其发生流行风险
棉花曲叶病的发生已被证实与介体烟粉虱种群丰度具有相关性(Ahmed et al.,2011),田间调查表明巴基斯坦棉花主产区旁遮普和信德的烟粉虱丰度分别高于经济阈值水平的15%和4%,且随着烟粉虱数量的增加,各省份的棉花曲叶病均显著增加(Ahmed et al.,2011)。室内传毒验证表明,25头印度本地隐种烟粉虱将CLCuMuV从朱槿传播到朱槿上的效率为20%,虫量增加至50头时,传播效率增加到70%(Rajeshwari et al.,2005),说明带毒虫量的高低直接影响朱槿曲叶病的流行程度。本研究中,烟粉虱优势种为传毒隐种AsiaⅡ7,但除广州市外,其他地区的潜在传毒烟粉虱隐种的数量均较少,各地烟粉虱虫株率大都在5.00%以下。推测这些地点CLCuMuV主要是随朱槿苗木的运输传入或烟粉虱携带病毒随苗木传入,而后经AsiaⅡ7烟粉虱小范围传播扩散。其他未调查到烟粉虱及烟粉虱优势隐种为MEAM1和MED的曲叶病发生地,可能是扦插感染病毒的朱槿枝条所致,具体情况需进一步研究。
虽然目前调查表明传毒隐种烟粉虱数量不多,但朱槿已作为一种重要的园林绿化植物在我国南方大面积种植(刘小冬等,2008),苗木跨区调运和种植苗木温室大棚的增加,可能会加速病毒和媒介烟粉虱从南方向内地扩散。因此,在朱槿盆栽花卉贸易及苗木跨区调运过程中应加强对CLCuMuV及烟粉虱的检测等相关措施,提早防范木尔坦棉花曲叶病毒病在我国棉区发生流行。
4 结论
我国多个地区的朱槿已感染CLCuMuV,木尔坦棉花曲叶病毒病在南方地区的广东、广西、海南、云南和福建5省(区)朱槿上发生严重。我国7省(区)中18个市(县)采样点朱槿上有烟粉虱发生,鉴定出的烟粉虱有5个隐种,其中AsiaⅡ7和MEAM1为优势种群。随着朱槿盆栽花卉贸易及苗木跨区调运,CLCuMuV有向内地逐渐扩散的趋势,应采取相应措施提早防范木尔坦棉花曲叶病毒在我国棉区发生流行。
致谢:新疆农业大学马德英教授、云南省农业科学院农业环境资源研究所谌爱东研究员、云南农业大学张晓明副教授、福建农林大学杜振国副教授、广西农业科学院植物保护研究所于永浩研究员等在样品收集中给予大力支持和帮助,谨此致谢!
参考文献:
陈婷,齐国君,石庆型,吕利华. 2019. 烟粉虱MEAM1和AsiaⅡ7隐种对不同寄主的选择性和生态适应性[J]. 环境昆虫学报,41(3):451-457. [Chen T,Qi G J,Shi Q X,Lü L H. 2019. Selectivity and adaptability between MEAM1 and AsiaⅡ7 cryptic species of Bemisia tabaci(Hemiptera:Aleyrodidae) to different host plants[J]. Journal of Environmental Entomology,41(3):451-457. ]
陈婷,齐国君,赵蕊,吕利华. 2016a. 广东朱槿病毒病发生及传播介体烟粉虱的种群组成调查初报[J]. 生物安全学报,25(3):204-208. [Chen T,Qi G J,Zhao R,Lü L H. 2016a. Incidence of leaf curl disease and the composition of Bemisia tabaci(Hemiptera:Aleyrodidae) population on Hibiscus rosa-sinensis in Guangdong Province,China[J]. Journal of Biosafety,25(3):204-208.]
陈婷,汤亚飞,赵蕊,何自福,吕利华. 2016b. 传播木尔坦棉花曲叶病毒的烟粉虱隐种鉴定[J]. 植物保护学报,43(1):91-98. [Chen T,Tang Y F,Zhao R,He Z F,Lü L H. 2016b. Identification of the cryptic of Bemisia tabaci transmitting cotton leaf curl Multan virus[J]. Journal of Plant Protection,43(1):91-98.]
董迪,何自福,柴兆祥. 2010. 广东黄秋葵黄脉曲叶病样中检测到烟粉虱传双生病毒[J]. 植物保护,36(1):65-68. [Dong D,He Z F,Chai Z X. 2010. Detection of whitefly-transmitted geminivirus associated with the okra yellow vein leaf curl disease in Guangdong[J]. Plant Protection,36(1):65-68.]
何自福,佘小漫,湯亚飞. 2012. 入侵我国的木尔坦棉花曲叶病毒及其为害[J]. 生物安全学报,21(2):87-92.[He Z F,She X M,Tang Y F. 2012. Cotton leaf curl Multan virus invading China and its damage potential[J]. Journal of Biosafety,21(2):87-92.]
林林,蔡健和,罗恩波,秦碧霞,胡冬梅,蒙姣荣,陈保善,农恒志. 2011. 南宁市朱槿曲叶病毒病病原分子鉴定和寄主范围研究[J]. 植物保护,37(4):44-47. [Lin L,Cai J H,Luo E B,Qin B X,Hu D M,Meng J R,Chen B S,Nong H Z. 2011. Molecular identification and host range of a geminivirus infecting Hibiscus in Nanning City[J]. Plant Protection,37(4):44-47.]
林莉,任顺祥,邱宝利. 2010. CV型烟粉虱在三种变叶木上的发育和繁殖[J]. 昆虫知识,47(6):1145-1149. [Lin L,Ren S X,Qiu B L. 2010. Effect of different variegated leafcroton to the growth and reproduction of ‘CVbiotype Bemisia tabaci[J]. Chinese Bulletin of Entomology,47(6):1145-1149 .]
刘小冬,姜卫兵,翁忙玲. 2008. 论木槿属树种及其在园林绿化中的应用[J]. 中国农学通报,24(8):315-320. [Liu X D,Jiang W B,Weng M L. 2008. On tree species of Hibiscus Linn and their application in landscape architecture[J]. Chinese Agricultural Science Bulletin,24(8):315-320.]
刘银泉,刘树生. 2012. 烟粉虱的分类地位及在中国的分布[J]. 生物安全学报,21(4):247-255. [Liu Y Q,Liu S S. 2012. Species status of Bemisia tabaci complex and their distributions in China[J]. Journal of Biosafety,21(4):247-255.]
毛明杰,何自福,虞皓,李华平. 2008. 侵染朱槿的木尔坦棉花曲叶病毒及其卫星DNA全基因组结构特征[J]. 病毒学报,24(1):64-68. [Mao M J,He Z H,Yu H,Li H P. 2008. Molecular characterization of Cotton leaf curl Multan virus and its satellite DNA that infects Hibiscus rosa-sinensis[J]. Chinese Journal of Virology,24(1):64-68.]
齐国君,陈婷,何自福,吕利华. 2016. 基于Maxent模型的棉花曲叶病在中国的适生性分析[J]. 棉花学报,28(5):443-451. [Qi G J,Chen T,He Z F,Lü L H. 2016. Maxent model-based analysis of the potential geographic distribution of cotton leaf curl disease in China[J]. Cotton Science,28(5):443-451.]
邱宝利,任顺祥,温硕洋,S. N. Mandour. 2006. 中国3种不同生物型烟粉虱的种群DNA多态性研究(英文)[J]. 华南农业大学学报,27(1):29-33. [Qiu B L,Ren S X,Wen S Y,Mandour S N. 2006. Population differentiation of three biotypes of Bemisia tabaci in China by DNA polymorphism[J]. Journal of South China Agricultural University,27(1):29-33.]
汤亚飞,何自福,杜振国,韩利芳,佘小漫,罗方芳. 2013. 侵染垂花悬铃花的木尔坦棉花曲叶病毒分子特征研究[J]. 植物病理学报,43(2):120-127. [Tang Y F,He Z F,Du Z G,Han L F,She X M,Luo F F. 2013. Molecular chara-cterization of the Cotton leaf curl Multan virus infecting Malvaiscus arboreus[J]. Acta Phytopathologica Sinica,43(2):120-127.]
汤亚飞,何自福,杜振国,佘小漫,蓝国兵. 2015a. 木尔坦棉花曲叶病毒及其伴随的β卫星分子复合侵染引起广东棉花曲叶病[J]. 中国农业科学,48(16):3166-3175. [Tang Y F,He Z F,Du Z G,She X M,Lan G B. 2015a. The complex of Cotton leaf curl Multan virus and its associated betasatellite molecule causing cotton leaf curl disease in Guangdong Province[J]. Scientia Agricultura Sinica,48(16):3166-3175.]
汤亚飞,何自福,杜振国,佘小漫,蓝国兵. 2015b. 海南红麻曲叶病的病原检测与鉴定[J]. 植物病理学报,45(6):561-568. [Tang Y F,He Z F,Du Z G,She X M,Lan G B. 2015b. Detection and identification of the pathogen cau-sing kenaf(Hibiscus cannabinus) leaf curl disease in Hainan Province of China[J]. Acta Phytopathologica Sinica,45(6):561-568.]
张晖,季英华,吴淑华,赵文浩,周彤,周益军. 2015. 江蘇朱槿上分离到的木尔坦棉花曲叶病毒基因组结构特征分析[J]. 植物病理学报,45(4):361-369. [Zhang H,Ji Y H,Wu S H,Zhao W H,Zhou T,Zhou Y J. 2015. Identification and characterization of Cotton leaf curl Multan virus from Hibiscus rosa-sinensis in Jiangsu[J]. Acta Phytopathologica Sinica,45(4):361-369.]
章松柏,夏宣喜,张洁,张友军,吴祖建. 2013. 福州市发生由木尔坦棉花曲叶病毒引起的朱槿曲叶病[J]. 植物保护,39(2):196-200. [Zhang S B,Xia X X,Zhang J,Zhang Y J,Wu Z J. 2013. A leaf curl disease on Hibiscus rose-sinensis in Fuzhou caused by Cotton leaf curl Multan virus[J]. Plant Protection,39(2):196-200.]
Ahmed M Z,de Barro P J,Greeff J M,Ren S X,Naveed M,Qiu B L. 2011. Genetic identity of the Bemisia tabaci species complex and association with high cotton leaf curl disease(CLCuD) incidence in Pakistan[J]. Pest Management Science,67(3):307-317.
Ashfaq M,Hebert P D N,Mirza M S,Khana M,Mansoor S,Shah G S,Zafar Y. 2014. DNA barcoding of Bemisia tabaci complex(Hemiptera:Aleyrodidae) reveals southerly expansion of the dominant whitefly species on cotton in Pakistan[J]. PLoS One,9(8):e104485.
Briddon R W,Bull S E,Amin I,Idris A M,Mansoor S,Bedford I D,Dhawan P,Rishi N,Siwatch S S,Abel-salam M A,Brown K J,Zafar Y,Marham G P. 2003. Diversity of DNA β:A satellite molecule associated with some monopartite begomoviruses[J]. Journal of Virology,312:106-121.
Briddon R W,Markham P G. 2000. Cotton leaf curl virus di-sease[J]. Virus Research,71(1/2):151-159.
Cai J H,Xie K,Lin L,Qin B X,Chen B S,Meng J R,Liu Y L. 2010. Cotton leaf curl Multan virus newly reported to be associated with cotton leaf curl disease in China[J]. Plant Pathology,59(4):794-795.
Chen T,Saeed Q,He Z F,Lü L H. 2019. Transmission efficiency of Cotton leaf curl Multan virus by three cryptic species of Bemisia tabaci complex in cotton cultivars[J].PeerJ,7:e7788.
Dinsdale A,Cook L,Rigion S C,Buckley Y M,de Barro P. 2010. Refined global analysis of Bemisia tabaci(Hemiptera:Sternorrhyncha:Aleyrodoidea:Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries[J]. Annals of the Entomological Socie-ty of America,103(2):196-208.
Du Z G,Tang Y F,He Z F,She X M. 2015. High genetic homogeneity points to a single introduction event responsible for invasion of Cotton leaf curl Multan virus and its associated betasatellite into China[J]. Virology Journal,12:163.
Frohlich D R,Torres-jerez I,Bedford I D,Markham P G,Brown J K. 1999. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers[J]. Molecular Ecology,8(10):1683-1691.
Liu S S,De Barro P J,Xu J,Luan J B,Zang L S,Ruan Y M,Wan F H. 2007. Asymmetric mating interactions drive widespread invasion and displacement in a whitefly[J]. Science,318:1769-1772.
Maharshi A,Yadav N K,Kumar P,Singh P,Singh J,Mukherjee A. 2017. An overview on cotton leaf curl disease:An emerging potential threat to cotton[J]. International Journal of Current Microbiology and Applied Sciences,6(10):2154-2162.
Mali V R. 1980. Studies on leaf curl disease of China rose in Marathwada[J]. Indian Journal of Mycology and Plant Pathology,9:119.
Mansoor S,Briddon R W,Bull S E,Bedford I D,Bashir A,Hussain M,Saeed M ,Zafar Y,Malik K A ,Fauquet C,Markhamp G. 2003. Cotton leaf curl disease is associated with multiple monopartite begomoviruses supported by single DNA β[J]. Archives of Virology,148(10):1969-1986.
Pan L L,Cui X Y,Chen Q F,Wang X W,Liu S S. 2018. Co-tton leaf curl disease:Which whitefly is the vector?[J]. Phytopathology,108(10):1172-1183.
Qiu B L,Dang F,Li S J,Ahmed M Z,Jin F L,Ren S X,Cuthbertsona G S. 2011. Comparison of biological parameters between the invasive B biotype and a new defined Cv biotype of Bemisia tabaci(Hemiptera:Aleyrodidae) in China[J]. Journal of Pest Science,84(4):419-427.
Rajeshwari R,Reddy R V C,Maruthi M N,Colvin J,Seal S E,Mumiyappa V. 2005. Host range,vector relationships and sequence comparison of a begomovirus infecting hibiscus in India[J]. Annals of Applied Biology,147(1):15-25.
Sattar M N,Kvarnheden V A,Saeed M,Briddon R W. 2013. Cotton leaf curl disease-an emerging threat to cotton production worldwide[J]. Journal of General Virology,94(4):695-710.
She X M,Tang Y F,He Z F,Lan G B. 2017. Molecular chara-cterization of Cotton leaf curl Multan virus and its associated betasatellite infecting Hibiscus rosa-sinensis in the Philippines[J]. Journal of plant pathology,99 (3):765-768.
Simon B,Cenis J L,Demichelis S,Rapisarda C,Caciagli P,Bosco D. 2003. Survey of Bemisia tabaci(Hemiptera:Aleyrodidae) biotypes in Italy with the description of a new biotype(T) from Euphorbia characias[J]. Bulletin of Entomological Research,93(2):259-264.
(責任编辑 麻小燕)
收稿日期:2020-09-23
基金项目:国家自然科学基金项目(31871937,32001973);广东省自然科学基金项目(2020A1515011098);广州市科技计划项目(201904010173)
作者简介:*为通讯作者:吕利华(1964-),研究员,主要从事农业害虫防治研究工作,E-mail:lhlu@gdppri.com;齐国君(1985-),副研究员,主要从事入侵生物防控研究工作,E-mail:qigj@gdppri.com。陈婷(1986-),主要从事入侵昆虫生物学研究工作,E-mail:chent@gdppri.com
