黄、东海渔业资源群落结构变化研究*
2020-02-13戴芳群陈云龙
戴芳群 朱 玲,2 陈云龙,2
黄、东海渔业资源群落结构变化研究*
戴芳群1朱 玲1,2陈云龙1,2①
(1. 农业农村部海洋渔业可持续发展重点实验室 山东省渔业资源与生态环境重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071;2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
为了解黄海和东海渔业资源群落结构的变化,基于2013年6月(夏季)和9月(秋季)黄、东海渔业资源底拖网调查数据,对渔业资源种类组成、优势种、渔获量空间分布、群落多样性和群落结构进行了研究。结果显示,共鉴定渔业种类185种,其中底层鱼类93种、中上层鱼类31种、甲壳类48种、头足类12种和贝类1种。在鱼类中,鲈形目种类最多,27科47属56种,占鱼类总种类数的45.2%。夏季优势种为鳀()、脊腹褐虾()、带鱼()和小黄鱼(),秋季优势种为三疣梭子蟹()、小黄鱼、鳀和带鱼,优势种季节波动不大。秋季平均单位时间渔获量为46.60 kg/h,高于夏季(39.35 kg/h)。秋季航次调查中渔业资源较夏季更为集中,主要分布在长江口和吕泗渔场附近,其他断面渔获量较低。夏季航次群落多样性指数高于秋季航次。CLUSTER聚类分析表明,黄、东海渔业生物群落可分为2组,夏、秋季2组界线变动不大,2组聚类的形成可能受水温和长江径流影响。
黄海;东海;渔业资源;群落结构;优势种
黄海作为全球50个大海洋生态系之一,位于中国大陆架边缘,渔业种类较为丰富,是我国重要的渔业水域(唐启升等, 2000)。黄海中部和南部海域是小黄鱼()、带鱼()、蓝点马鲛()和鳀()等多种重要经济鱼类的产卵场和索饵场,也是洄游性鱼类进出渤海的通道,形成了长江口渔场、吕泗渔场、海州湾渔场和烟威渔场,为我国及周边国家提供了丰富的渔业资源(Jin, 2003)。黄海海洋环境复杂,中部海域水深变化剧烈,底质类型多为基岩岸与砂砾质;而黄海南部受古黄河冲击,底质类型多为淤泥粉砂质,形成了苏北浅滩及独特的辐射沙洲地形(肖欢欢等, 2017)。黄海中部和南部海区受黄海冷水团、海州湾和吕泗海区的上升流、苏北沿岸流和长江冲淡水影响,海洋环境因子区域变化明显,形成了生境差异较大的不同断面,造成了黄海中部和南部鱼类群落不同的生物地理分布格局(于振海等, 2010; 单秀娟等, 2016)。东海位于中国大陆中部东侧,北连黄海,长江、钱塘江等多条河流注入东海,带来丰富的营养盐,东海受黑潮暖流及其西侧分支-台湾暖流、长江冲淡水及黄海冷水团的综合作用影响,基础饵料丰富,在河口附近的浅海海域为主要经济鱼类的产卵场,在沿岸或近海海域分布着多种经济鱼类的索饵场所,东海近海的外侧和南部水域又是多种经济鱼类的越冬场,东海是中国重要的天然渔场,也是世界上高生产力水域之一(姜亚洲等, 2008)。
国内学者对该区域的研究主要集中在黄、东海及长江口海域鱼类群体结构(刘勇等, 2006; 单秀娟等, 2014; 汤昌盛等, 2017),而对黄海和东海海域整个渔业资源群落结构的季节变动研究较少(单秀娟等, 2013)。2003~2017年,35°N以北的黄海和渤海海域休渔期为7月1日~9月1日,26°30¢N~35°N的黄海和东海海域休渔期为6月16日~9月15日(郑元甲等, 2003),6月和9月的调查资料更能代表黄海南部和东海海域休渔期早期和后期的资源量水平,对于评估伏季休渔制度对资源的养护效果具有重要的基础意义。
1 材料与方法
1.1 材料来源与分析
样品取自2013年6月(夏季)和9月(秋季)在黄、东海进行的渔业生物底拖网调查。调查区域为27°24¢59²~37°00¢00²N,120°30¢00²~124°30¢00²E,以与等深线近乎垂直的方式布设了11条大致等间隔的断面,共计47个站位(图1)。调查船为中国水产科学研究院黄海水产研究所“北斗”号,其网具参数为网目836目´20 cm,网口周长为167.2 m,网衣长度为83.2 m,囊网网目为24 mm,拖速为3 kn,每站拖网1 h左右。在实验室内对渔获生物进行种类鉴定和生物学测定,记录每一种的重量和尾数,计算单位时间渔获量(kg/h)和单位时间渔获尾数(ind./h)。
1.2 相对重要性指数
相对重要性指数(Index of relative importance,IRI)(Pianka, 1971)计算公式如下:
IRI=(%+%)×%
式中,%为某一种类的尾数占总尾数的百分比;%为某一种类的重量占总重量的百分比;%为某一种类出现的站数占调查总站数的百分比。根据程济生等(2000),当IRI>500时,该种为优势种,500~100为常见种,100~10为一般种,10~1为少见种,IRI<1为稀有种。
1.3 群落多样性指数
不同种类及同种类个体大小差异很大,用生物量表示的多样性更接近种类间能量分布,因此,本研究根据生物量计算渔业生物群落种类多样性。多样性计算方法如下:

图1 黄、东海渔业资源调查站位
(1) Margalef的种类丰富度指数(Margalef, 1958):
=(-1)/ln
式中,为总种数,为总渔获量。
(2) Shannon-Wiener多样性指数(Shannon, 1949):
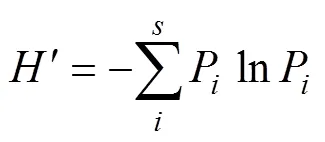
(3) Pielou均匀度指数(Pielou, 1975):
¢¢/ln
式中,¢为种类均匀度,为总种数。
1.4 群落结构分析
应用未加权的组平均方法(UPGMA)进行聚类(CLUSTER)分析,使用单因子相似性分析(ANOSIM)对不同组群落结构差异进行显著性检验,并利用相似性百分比分析(SIMPER)分析造成各组内群落结构相似的典型种以及造成不同组之间群落结构差异的分歧种(Clarke, 1993)。用Bray-Curtis相似性指数计算不同季节间渔业资源群落结构组成的差异(Field,1982):


以上数据统计分析采用软件PRIMER5完成。
2 结果
2.1 渔业资源种类组成和优势种季节变化
2013年夏季和秋季调查共捕获渔业种类185种,隶属于25目98科148属,其中底层鱼类93种、中上层鱼类31种、头足类12种、甲壳类48种和贝类1种,分别占总种数的50.3%、16.8%、6.5%、25.9%和0.5%。鱼类中,鲈形目种类最多,27科47属 56种,占鱼类总种类数的45.2%,其次为鲉形目, 10科14属16种,其余各目均在9种以下;甲壳类共有3目22科32属48种,主要为十足目种类;头足类3目6科9属12种(表1)。夏季调查种类共计 149种,其中底层鱼类74种、中上层鱼类21种、头足类42种、甲壳类11种和贝类1种,分别占总种数的49.7%、14.1%、28.2%、7.4%和0.7%(表2)。秋季调查种类共计143种,底层鱼类71种、中上层鱼类23种、头足类39种和甲壳类10种,分别占总种数的49.7%、16.1%、27.3%和7.0% (表2)。
表1 2013年夏季和秋季黄、东海渔业种类各分类阶元的组成

Tab.1 Taxonomic category of fishery resources in the Yellow Sea and East China Sea during summer and autumn in 2013
表2 2013年夏季和秋季黄、东海各生态类型种类统计

Tab.2 Species composition of different fishery ecotypes in the Yellow Sea and East China Sea during summer and autumn in 2013
根据相对重要性指数,夏季渔获优势种为鳀、脊腹褐虾()、带鱼和小黄鱼(表3),4个优势种平均单位时间渔获量分别为13.33、7.73、14.76和2.06 kg/h,这4个优势种占夏季总渔获量和渔获尾数的比例分别为51.4%和56.7%,其中,鳀、脊腹褐虾、小黄鱼和带鱼分别占总渔获量的20.9%、9.2%、17.6%和3.7%。秋季优势种为三疣梭子蟹()小黄鱼、鳀和带鱼,其平均单位时间渔获量为17.76、7.55、5.99和20.25 kg/h,分别占总渔获量的19.5%、11.6%、7.6%和20.0%。夏季和秋季优势种主要由底层鱼类和甲壳类组成,中上层鱼类仅出现了鳀。小黄鱼、带鱼和鳀均为夏季和秋季的优势种,3种合计占夏季和秋季调查渔获量比例分别为42.2%和39.2%。
表3 2013年夏季和秋季黄、东海渔业资源优势种季节变化

Tab.3 Seasonal variations of dominant fishery resources during summer and autumn in 2013
2.2 渔获量空间分布和季节变化
秋季平均单位时间渔获量为46.60 kg/h,高于夏季(39.35 kg/h)。夏季和秋季渔获中,底层鱼类和中上层鱼类平均单位时间渔获量较高。夏季调查中,底层鱼类和中上层鱼类平均单位时间渔获量分别为17.27 kg/h和10.68 kg/h,其渔获量占总渔获量的百分比分别为43.88%和27.15%。秋季调查中,底层鱼类和中上层鱼类平均单位时间渔获量分别为23.43和11.40 kg/h,其渔获量分别占总渔获量的50.27%和24.46%。2个季节中,头足类渔获量较低,平均单位时间渔获量分别为1.21和0.48 kg/h。
夏季调查东海断面中部站位渔获量百分比较高,黄海中部冷水团位置渔获量百分比均较低;秋季渔业资源较夏季更为集中,主要分布在长江口和吕泗渔场附近位置,平均单位时间渔获量最高(178.51 kg/h),其他断面渔获量较低(图2)。
2.3 群落多样性特征及其分布
夏季和秋季调查渔获种类数分别为149种和143种。夏季总丰富度指数和总多样性总指数均最大,各站位分别在0.16~4.43和0.33~2.36之间波动;秋季丰富度指数和多样性总指数较小,各站位分别在0~3.87和0~2.33之间波动,夏季平均丰富度指数(1.99)较秋季(1.69)高(表4)。在夏季调查中,多样性指数最高站位出现在东海北部浙江近岸站位,苏北浅滩区2个断面近岸站位多样性指数较高,而长江口外断面站位多样性指数较低(图4)。秋季调查中,长江口区域断面中部和外部站位多样性指数较高,其他断面多样性指数偏低。以重量计算的相似性指数,夏季和秋季的相似性指数为0.54;以尾数计算的相似性指数,夏季和秋季的相似性指数为0.30。

图2 2013年夏季和秋季黄、东海渔业资源平均单位时间渔获量的空间分布
表4 2013年夏季和秋季黄、东海渔业种类多样性指数的变化

Tab.4 Variations of fishery species diversity indices during summer and autumn in 2013

图3 2013年夏季和秋季黄、东海渔业生物多样性指数H¢、D和J¢的空间分布
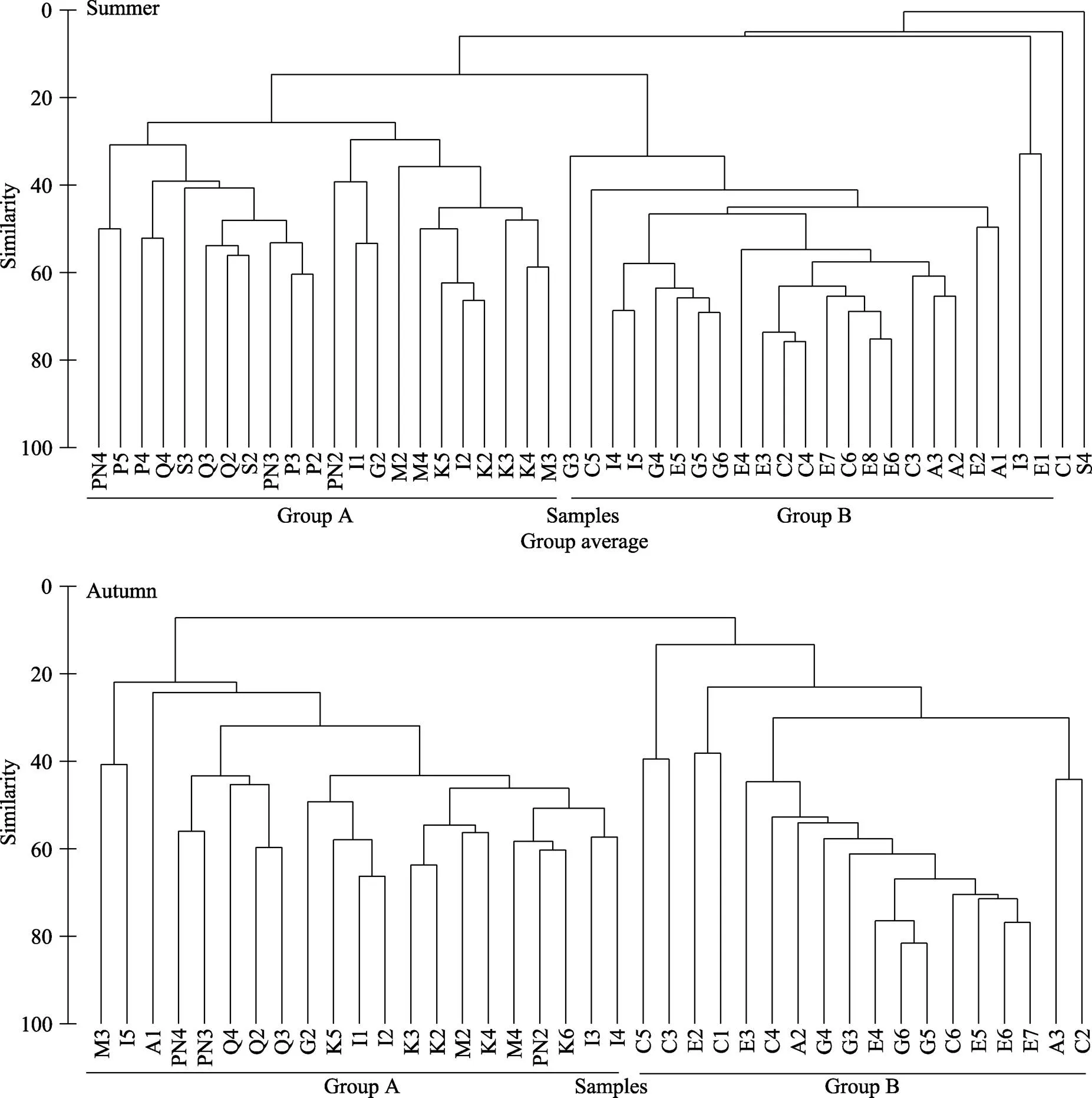
图4 2013年夏季和秋季黄、东海渔业生物群落聚类分析
2.4 群落结构分析


图5 2013年夏季和秋季黄、东海渔业资源群落结构划分示意
3 讨论
在本调查中,共鉴定渔业种类185种,从物种组成看,渔获物以底层鱼类和甲壳类为主,该结果同长江口区域、黄海南部区域和浙江近岸海域的调查结果相符(金显仕等, 2009; 单秀娟等, 2013; 孙鹏飞等, 2015; 汤昌盛等, 2017)。单秀娟等(2013)对山东以南黄海海域的4个季度渔业资源调查结果进行统计,鉴定出渔获种类156种,其中底层鱼97种,甲壳类为36种。近20年来,对长江口及其邻近海域渔业资源调查物种统计结果显示,底层鱼类和甲壳类占优势地位。根据李建生等(2004)的研究,2000~2002年长江口海域各季节均以鱼类为绝对优势类群,甲壳类春季所占百分比最高,为11.1%。而孙鹏飞等(2015)研究显示,2012~2013年,在长江口区域甲壳类在渔获中所占比例为40.1%~54.2%。本研究中,甲壳类物种数占总渔获种类数的25.9%。比较不同年代黄、东海渔业资源调查研究结果同本研究发现,甲壳类在黄海南部区域物种数和资源量逐渐增加。甲壳类在渔业生态系统中处于承上启下的位置,高强度的捕捞致使传统营养级较高的经济鱼类资源量下降,甲壳类被捕食的压力减小,同时,近海富营养化为营养级较低的甲壳类资源量的增加提供了物质基础,渔业生物中甲壳类种类数及资源量相对增加(李涛等, 2011)。从鱼类物种组成看,调查各断面鱼类种类以鲈形目最多,其次是鲉形目,鱼类主要由暖温种和暖水种组成,仅大头鳕、玉筋鱼()等少量冷温种,同单秀娟等(2013)对黄海中南部鱼类种类组成研究结果一致。各断面渔获种类组成存在明显的夏秋季节变化,这与鱼类、甲壳类和头足类季节间产卵、索饵及越冬洄游有关(张波等, 2009; 杨涛等, 2018)。


秋季平均单位时间渔获量明显高于夏季,可能与伏季休渔政策有关,经过3个月休渔,渔业生物幼体个体数量增加,并且渔业生物得到充分生长,体重也明显增加,夏季调查中,A、C、K、M断面的中部站位,断面G、I、P、Q、S近岸站位渔获量较高,各断面外海站位普遍偏低。秋季调查中,渔获量向长江口,北部断面渔获量较低。这可能与鳀、小黄鱼、带鱼、银鲳和三疣梭子蟹等越冬场鱼群的洄游有关,这些鱼类随沿岸水温升高5~6月在长江口和吕泗渔场附近形成产卵群体或索饵群体,6月以后,索饵群体北上山东半岛南部海域索饵,9月以后逐渐回到长江口及其邻近海域,一部分群体游向外海越冬场,一部分群体继续南下(宋海棠等, 1989; 邓景耀等, 1991; 水柏年, 1999; 郑元甲等, 2003;徐兆礼等, 2009)。
黄海中部、南部和东海北部的渔业生物群落夏季多样性指数、均匀度指数和丰富度指数均高于秋季。夏秋季群落丰富度指数和多样性指数均显著低于2006年的黄海对应季节的研究结果(2.79~6.20;¢0.78~3.05),均匀度指数同2006年的调查结果一致 (单秀娟等, 2013)。夏季长江口附近的K、M、PN 3个断面近岸和中部站位群落多样性指数较低,这可能是由于夏季长江径流较强,冲淡水影响较广,造成该区域近海种类偏低。随着长江径流减少,秋季长江附近的K、M、PN 3个断面物种多样性明显升高。
聚类分析结果表明,黄海中部、南部和东海北部海域渔业生物群落划分为2个地理组,分界线在断面I,夏秋季2组界线变动不大。2组聚类反映了黄海不同区域鱼类适温类型的差异。A组包括黄海南部和东海北部水域,该组以暖水性和暖温性种类为主,并且多为中上层小型种类,A组站位主要受长江径流影响,营养盐较丰富,初级生产力较高。B组包括黄海中部的4个断面,主要以暖温种类为主,存在一定量的冷温性种类,受黄海冷水团影响。黄海冷水团为夏季冷温性种类提供了庇护场所,限制了部分底层暖温性种类的向北扩散。本研究聚类结果与单秀娟等(2014)对黄海中南部鱼类群落结构划分一致。聚类结果支持在深度梯度变化不大的温带水域的鱼类群落组成在很大程度上存在温度依赖性的结论(程济生等, 2004)。除温度外,深度也是影响渔业资源群落的重要因素,东海大陆架鱼类群落的种类组成沿着水深梯度变化明显,每种鱼类均有一定的水深范围(李圣法等, 2007)。在本研究中,由于南北温度变化尺度较大,深度梯度变化区域不明显,因此,在深度梯度变化不明显的情况下鱼类群聚主要与水温相关。
Cheng JS, Yu LF. The change of structure and diversity of demersal fish communities in the Yellow Sea and East China Sea in winter. Journal of Fisheries of China, 2004, 28(1): 29–34 [程济生, 俞连福. 黄、东海冬季底层鱼类群落结构及多样性变化. 水产学报, 2004, 28(1): 29–34]
Cheng JS. The structure and diversity of demersal fish communities in winter in the East China Sea and the Yellow Sea. Marine Fisheries Research, 2000, 21(3): 1–8 [程济生. 东、黄海冬季底层鱼类群落结构及其多样性. 海洋水产研究, 2000, 21(3): 1–8]
Clarke KR, Ainsworth M. A method of linking multivariate community structure to environmental variables. Marine Ecology-Progress Series, 1993, 92(3): 205–219
Deng JY, Zhao CY. Marine fishery biology. Beijing: China Agriculture Press, 1991 [邓景耀, 赵传絪. 海洋渔业生物学. 北京: 农业出版社, 1991]
Field JG, Clarke KR, Warwick RM. A practical strategy for analysing multispecies distribution patterns. Marine Ecology- Progress Series, 1982, 8: 37–52
Jiang YZ, Cheng JH, Li SF. Variation in fish community structure and biodiversity in the north of the East China Sea between two periods. Journal of Fishery Sciences of China, 2008, 15(3): 453–459 [姜亚洲, 程家骅, 李圣法. 东海北部鱼类群落多样性及其结构特征的变化. 中国水产科学, 2008, 15(3): 453–459]
Jin XS, Xu B, Tang QS. Fish assemblage structure in the East China Sea and southern Yellow Sea during autumn and spring. Journal of Fish Biology, 2003, 62(5): 1194–1205
Jin XS, Shan XJ, Guo XW,. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters. Acta Ecologica Sinica, 2009, 29(9): 4761–4772 [金显仕, 单秀娟, 郭学武, 等. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 2009, 29(9): 4761–4772]
Jin XS. The dynamics of major fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 2000, 7(4): 22–26 [金显仕. 渤海主要渔业生物资源变动的研究. 中国水产科学, 2000, 7(4): 22–26]
Li JS, Li SF, Ren YP,. Seasonal variety of fishery biology community structure in fishing ground of the Yangtze estuary. Journal of Fishery Sciences of China, 2004, 11(5): 432–439 [李建生, 李圣法, 任一平, 等. 长江口渔场渔业生物群落结构的季节变化. 中国水产科学, 2004, 11(5): 432–439]
Li SF, Cheng JH, Yan LP. Spatial structures of fish communities on the continental shelf of the East China Sea. Acta Ecologica Sinica, 2007, 27(11): 4377–4386 [李圣法, 程家骅, 严利平. 东海大陆架鱼类群落的空间结构. 生态学报, 2007, 27(11): 4377–4386]
Li T, Zhang XM, Zhang PD,. Seasonal variation on community structure of fishery resources in the coastal waters of southern Shandong Peninsula. Periodical of Ocean University of China, 2011, 41(1/2): 41–50 [李涛, 张秀梅, 张沛东, 等. 山东半岛南部近岸海域渔业资源群落结构的季节变化. 中国海洋大学学报, 2011, 41(1/2): 41–50]
Liu Y, Li SF, Cheng JH. A study on seasonal changes of the fish communities in the East China Sea and the Huanghai Sea. Acta Oceanologica Sinica, 2006, 28(4): 108–114 [刘勇, 李圣法, 程家骅. 东海、黄海鱼类群落结构的季节变化研究. 海洋学报, 2006, 28(4): 108–114]
Margalef R. Information theory in ecology. General Systems, 1958, 3: 36–71
Pianka ER. Ecology of the agamid lizardin Western Australia. Copeia, 1971(3): 527–536
Pielou EC. Ecological diversity. New York: John Wiley and Sons, 1975
Shan XJ, Chen YL, Dai FQ,. Variations in fish community structure and diversity in the sections of the central and southern Yellow Sea. Acta Ecologica Sinica, 2014, 34(2): 377–389 [单秀娟, 陈云龙, 戴芳群, 等. 黄海中南部不同断面鱼类群落结构及其多样性. 生态学报, 2014, 34(2): 377–389]
Shan XJ, Chen YL, Jin XS,. Reshaping the distribution of fish abundance by climate change: A case study in the Yangtze River estuary. Progress in Fishery Sciences, 2016, 37(6): 1–10 [单秀娟, 陈云龙, 金显仕, 等. 气候变化对长江口鱼类资源密度分布的重塑作用. 渔业科学进展, 2016, 37(6): 1–10]
Shan XJ, Sun PF, Jin XS,. Seasonal variations of fishery resource structure in the sections of the southern Yellow Sea. Journal of Fisheries of China, 2013, 37(3): 425–435 [单秀娟, 孙鹏飞, 金显仕, 等. 黄海典型断面渔业资源结构的季节变化. 水产学报, 2013, 37(3): 425–435]
Shannon CE, Weaver W. The mathematical theory of communication. Urbana: University of Illinois Press, 1949
Shui BN. Preliminary study on the distribution ofand fishing grounds in the Yellow Sea and East China Sea. Journal of Fisheries of Zhoushan, 1999, S(57): 17–20 [水柏年. 黄、东海鳀鱼洄游及渔场分布初探. 舟山渔业, 1999, S(57): 17–20]
Song HT, Ding YP, Xu YJ. Migration and population composition ofin the northern Zhejiang Province. Marine Science Bulletin, 1989, 8(1): 66–74 [宋海棠, 丁耀平, 许源剑. 浙北近海三疣梭子蟹洄游分布和群体组成特征. 海洋通报, 1989, 8(1): 66–74]
Sun PF, Dai FQ, Chen YL,. Seasonal variations in structure of fishery resource in the Yangtze River estuary and its adjacent waters. Progress in Fishery Sciences, 2015, 36(6): 8–16 [孙鹏飞, 戴芳群, 陈云龙, 等. 长江口及其邻近海域渔业资源结构的季节变化. 渔业科学进展, 2015, 36(6): 8–16]
Tang CS, Zhang F, Feng S,. Biological community of fishery resources in the Yangtze River estuary and adjacent sea areas in the summer of 2015. Marine Fisheries, 2017, 39(5): 490–499 [汤昌盛, 张芳, 冯颂, 等. 2015年夏季长江口及其邻近海域渔业生物群落结构分析. 海洋渔业, 2017, 39(5): 490–499]
Tang QS, Su JL. Marine ecosystem dynamics research of China I. Key scientific issues and research development strategies. Beijing: Science Press, 2000 [唐启升, 苏纪兰. 中国海洋生态系统动力学研究Ⅰ. 关键科学问题与研究发展战略. 北京: 科学出版社, 2000]
Xiao HH, Zhang CL, Xu BD,. Spatial pattern of ichthyoplankton assemblage in the coastal waters of central and southern Yellow Sea in the spring. Haiyang Xuebao, 2017, 39(8): 34–47 [肖欢欢, 张崇良, 徐宾铎, 等. 黄海中南部近岸海域春季鱼类浮游生物群落空间格局研究. 海洋学报, 2017, 39(8): 34–47]
Xu BD, Jin XS, Liang ZL. Changes of demersal fish community structure in the Yellow Sea during the autumn. Journal of Fishery Sciences of China, 2003, 10(2): 148–154 [徐宾铎, 金显仕, 梁振林. 秋季黄海底层鱼类群落结构的变化. 中国水产科学, 2003, 10(2): 148–154]
Xu ZL, Chen JJ. Analysis on migratory routine of. Journal of Fishery Sciences of China, 2009,16(6): 931–940 [徐兆礼, 陈佳杰. 小黄鱼洄游路线分析. 中国水产科学, 2009, 16(6): 931–940]
Yang T, Shan XJ, Jin XS,. Long-term changes in keystone species in fish community in spring in Laizhou Bay. Progress in Fishery Sciences, 2018, 39(1): 1–11 [杨涛, 单秀娟, 金显仕, 等. 莱州湾春季鱼类群落关键种的长期变化. 渔业科学进展, 2018, 39(1): 1–11]
Yu ZH, Jin XS, Li XS. Analysis of ecological niche for major fish species in the central and southern Yellow Sea. Progress in Fishery Sciences, 2010, 31(6): 1–8 [于振海, 金显仕, 李显森. 黄海中南部主要鱼种的生态位分析. 渔业科学进展, 2010, 31(6): 1–8]
Zhang B, Tang QS, Jin XS. Functional groups of communities and their major species at high trophic level in the Yellow Sea ecosystem. Acta Ecologica Sinica, 2009, 29(3): 1099– 1111 [张波, 唐启升, 金显仕. 黄海生态系统高营养层次生物群落功能群及其主要种类. 生态学报, 2009, 29(3): 1099–1111]
Zhang B, Tang QS. Study on trophic level of important resources species at high troph levels in the Bohai Sea, Yellow Sea and East China Sea. Advances in Marine Science, 2004, 22(4): 393–404 [张波, 唐启升. 渤、黄、东海高营养层次重要生物资源种类的营养级研究. 海洋科学进展, 2004, 22(4): 393–404]
Zheng YJ, Chen XZ, Cheng JH,. East China Sea continental shelf biological resources and environment. Shanghai: Shanghai Science and Technology Press, 2003 [郑元甲, 陈雪忠, 程家骅, 等. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 2003]
Variations of Fishery Resource Structure in the Yellow Sea and East China Sea
DAI Fangqun1, ZHU Ling1,2, CHEN Yunlong1,2①
(1. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs; Shandong Provincial Key Laboratory of Fishery Resources and Eco-Environment, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071;2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071)
To elucidate the dynamics of fishery resources structure in the Yellow Sea and East China Sea, species composition, dominate species, spatial distribution of catch, community diversity and community structure of fishery species in the Yellow Sea and East China Sea were studied based on the bottom trawl survey data collected from June and September in 2013. A total of 185 fishery species were collected, including 93 dermersal fish species, 31 pelagic species, 48 crustacean species, 12 cephalopod species and one mollusk species. Among these fish species, the richest species was found in Perciformes (27 families, 47 genera and 56 species). There was no obvious seasonal variation of dominate species. In summer, the dominate species were sand shrimpand hairtail, whereas in autumn, the dominate species were the swimming crab, yellow small croaker, Japanese anchovyand hairtail. The average catch per haul was higher in autumn (46.60 kg/h) than that in summer (39.35 kg/h). The spatial distribution of fishery resource in autumn concentrated in the Yangtze River estuary and Lvsi fishing ground. The diversity indices of fishery species in summer were higher than that in autumn. The fishery biological communities could be divided into two groups based on the cluster analysis. The separation between two groups was primarily influenced by the temperature and discharge of Yangtze River.
Yellow Sea; East China Sea; Fishery resources; Community structure; Dominate species
S931
A
2095-9869(2020)01-0001-10
10.19663/j.issn2095-9869.20181120001
* 青岛海洋科学与技术国家实验室“鳌山人才”培养计划项目(2017ASTCP-ES07)、国家重点基础研究发展计划(2017YFC1404402)和山东省泰山学者专项基金项目共同资助[This work was supported by Aoshan Talents Cultivation Program Supported by Qingdao National Laboratory for Marine Science and Technology (2017ASTCP-ES07), the National Key Research and Development Program of China (2017YFC1404402), and Special Funds for Taishan Scholar Project of Shandong Province]. 戴芳群,E-mail: dai@ysfri.ac.cn
陈云龙,E-mail: chenyl@ysfri.ac.cn
2018-11-20,
2018-12-20
http://www.yykxjz.cn/
戴芳群, 朱玲, 陈云龙. 黄、东海渔业资源群落结构变化研究. 渔业科学进展, 2020, 41(1): 01–10
Dai FQ, Zhu L, Chen YL. Variations of fishery resource structure in the Yellow Sea and East China Sea. Progress in Fishery Sciences, 2020, 41(1): 01–10
CHEN Yunlong, Email: chenyl@ysfri.ac.cn
(编辑 马璀艳)
