水分胁迫对极小种群东兴金花茶幼苗光合特性的影响
2020-01-18唐健民柴胜丰邹蓉陈宗游史艳财蒋运生韦霄
唐健民 柴胜丰 邹蓉 陈宗游 史艳财 蒋运生 韦霄


摘 要:為了解东兴金花茶幼苗对水分胁迫的适应能力和响应机制,该文以东兴金花茶1年生实生苗为材料,采用盆栽控水试验,研究不同控水时间处理对东兴金花茶幼苗的生理生态特性的影响。结果表明:随着控水时间的延长,水分胁迫的程度不断加剧,东兴金花茶叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)呈现显著下降趋势;胞间CO2浓度(Ci)呈现先低后高的变化趋势,水分利用效率(WUE)呈现先高后低的变化趋势。土壤含水率和叶片相对含水量均呈现不断下降的趋势,丙二醛呈现先降后升的变化趋势;东兴金花茶幼苗的荧光参数Fv/Fm和Fv/Fo呈现先增加后不断下降趋势,分别从0.806下降至0.754和4.17下降至3.08,表明水分胁迫降低了PS Ⅱ原初光能转化效率,光合作用原初反应过程受到抑制。基于水分胁迫的生理生态指标和叶片生物性状的变化表明,控水时间在4 d情况下东兴金花茶可以提高自身水分利用效率来抵抗干旱,说明东兴金花茶幼苗对水分胁迫具有一定的适应性和响应机制。控水8~12 d,东兴金花茶的光合指标下降显著,土壤含水率下降至14.157%~15.065%,其叶片萎蔫、打卷,低于此水平东兴金花茶幼苗会因过度干旱而死亡,表明东兴金花茶幼苗对水分胁迫的耐受极限土壤含水率为14.157%~15.065%。研究结果有助于营建适宜的环境以保证东兴金花茶的正常生长和繁殖,对东兴金花茶迁地保护、引种培育和回归自然具有重要的科学指导意义。
关键词:东兴金花茶, 不同控水时间, 光合作用, 叶绿素荧光参数, 种群扩散
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2020)12-1764-09
Abstract:In order to understand the adaptability and response mechanism of Camellia tunghinensis seedlings to water stress, a pot experiment was conducted to study the effects of different water control time on the physiological and ecological characteristics of C. tunghinensis seedlings. The results were as follows:Net photosynthetic rate(Pn), stomatal conductance(Gs) and transpiration rate(Tr) decreased significantly with the extension of water control time and the aggravation of water stress; The variation trend of intercellular CO2 concentration(Ci)was low first and then high, and water use efficiency (WUE) was high first and then low. Soil water content and leaf relative water content showed a decreasing trend, malondialdehyde first decreased and then increased; The fluorescence parameters Fv/Fm and Fv/Fo of C. tunghinensis seedlings first increased and then decreased continuously, from 0.806 to 0.754 and 4.17 to 3.08, respectively, which showed that water stress reduced PS Ⅱprimary light energy conversion efficiency, impaired the potential activity of PS Ⅱ, and inhibited the primary reaction process of photosynthesis. Based on the above changes of physiological and ecological indexes of water stress and biological characters of leaves, C. tunghinensis could improve its own water use efficiency to resist drought in the case of water control time of 4 d, indicating that seedlings had certain adaptability and response mechanism to water stress. In 8-12 d, the photosynthetic indexes of C. tunghinensis decreased significantly, and soil water content decreased to 14.157%-15.065%, which leaf wilting, curl, and lower than this level seedlings will die due to excessive drought, indicating that the tolerance limit of C. tunghinensis seedlings to water stress was 14.157%-15.065% of soil water content. The results of the study are helpful to the establishment of a suitable environment to ensure the normal growth and reproduction of C. tunghinensis, which is of important scientific significance to the conservation, cultivation and return to nature.
Key words:Camellia tunghinensis, different water control time, photosynthesis, chlorophyll fluorescence parameters, population dispersion
东兴金花茶(Camellia tunghinensis),金花茶组(Sect . Chrysantha Chang)的常绿灌木或小乔木,广西特有的国家二级保护植物(黄仕训等,2006)。随着金花茶组植物知名度不断提高,种植、研究、开发金花茶组植物成为热潮(杨期和等,2010),导致野生金花茶组植物遭受到掠夺性开采,造成在过去的30年中金花茶的野生资源消失殆尽,东兴金花茶作为珍贵稀有的金花茶资源,处于一种岌岌可危的灾难性状态(刘林,1999)。大力开展东兴金花茶保护生物学研究,摸清其濒危机制,制定科学合理的保护策略,势在必行。
目前,学者们已对东兴金花茶的遗传多样性(唐健民等,2014)、种子繁殖(杨泉光等,2012)和扦插繁殖(蒋运生等,2010)等多领域开展了研究。其中唐健民等(2014)研究显示东兴金花茶的种群遗传多样性较高及杂合子有过剩现象;杨泉光等(2012)和蒋运生等(2010)研究表明东兴金花茶的种子及其无性扦插繁育都具有较高的萌发率(70%)和成活率(80%)。上述结果都表明东兴金花茶的繁殖系统可能不是限制种群发展的主要原因。因此,限制东兴金花茶种群难以扩散的原因有可能与其生存环境及本身对光、水的需求相关。东兴金花茶在自然环境中仅分布于十万大山南坡的上岳保护区。十万大山主脉由东向西横贯中部,是广西南部重要的气候分界线,南坡迎风,属南亚热带湿热季风气候,雨水丰富,干湿季分明;北坡背风,受北部湾海洋气候影响,常出现冬春旱灾,南北坡气候差异较大。光照和水分因素对植物生长影响非常广泛且深刻,它影响着植物各个生长发育阶段,对植物生存、生长和发育非常重要(吴富勤等,2015;胡小京等,2020)。植物对环境的适应取决于生存环境中水分的供应状况,多数濒危植物在水分和光合生态方面存在适应力、生存力较差的共性,表现出不耐干旱的生理特性,干旱成为大多数濒危植物种群分布小、难以扩散的主要限制因素(邓云等,2008;赵丽丽等,2019)。
针对东兴金花茶种群分布狭小、难以扩散的原因,开展不同水分胁迫对东兴金花茶光合特性的影响,通过不同的控水时间处理,模拟自然环境下土壤蒸发及叶片的蒸腾作用等水分散失途径,形成不同程度的干旱胁迫水平,研究短期内东兴金花茶对水分胁迫的耐受性,揭示不同干旱状态下东兴金花茶光合特性的变化规律和响应机制,分析其对干旱环境适应能力及适应机制,尋找限制东兴金花茶幼苗生长发育的土壤水分条件,为后续东兴金花茶的生态自然回归、扩大种群规模提供理论基础和技术指导。
1 材料与方法
1.1 试验地概况
试验地设在广西壮族自治区中国科学院广西植物研究所(110°07′ E、25°11′ N)金花茶种质圃内进行。桂林属中亚热带季风性气候区,年均气温19.2 ℃,有6~7个月的月平均气温高于20 ℃,年相对湿度78.0%,干湿季明显。
1.2 材料和方法
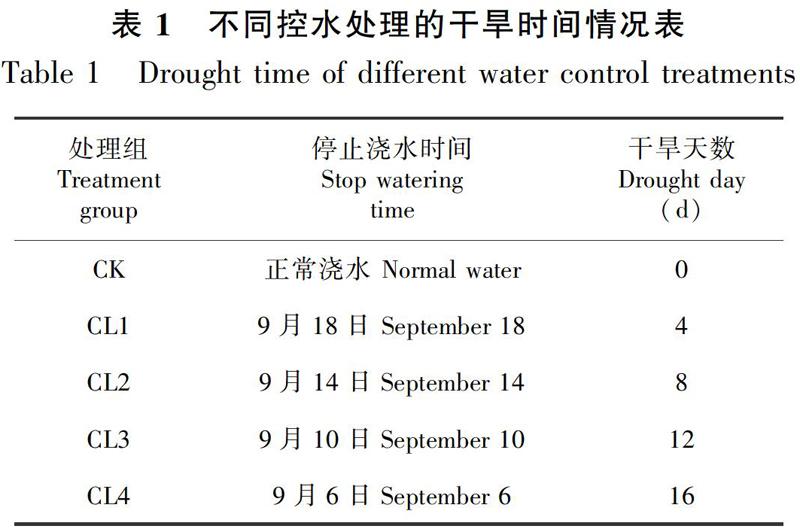
于6月精选东兴金花茶健康、长势一致及任何无病虫害的当年生幼苗,将幼苗移栽至塑料花盆中,基质为黄土,光照系数为全光照(玻璃房内的光强是自然光强的25%左右,与东兴金花茶野外生境相似,位于茂密的乔木层下,郁闭度在0.75~0.8之间),温度设为室温。每天固定时间进行浇水一次(以灌满花盆为标准),确保每株苗木与土壤中影响干旱的因子一致。通过适应性阶段后,9月2日开始对每组进行控水处理试验(表1),试验分成5组,待CL1停止浇水3 d后,即9月22日测定不同水分胁迫程度的各项生理生态指标。
1.3 测定项目与方法
所有控水处理都达到设定的水分胁迫时间,分别选择东兴金花茶植株顶端向下成熟叶片进行光合测定。然后分别选择各不同控水时间中长势接近的成熟叶片,剪碎后分装用于测定土壤含水率、叶片相对含水量、丙二醛、光合色素等各项指标。
光合测定:采用Li-6400 光合仪进行测定,测定指标主要有净光合速率光合(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等光合指标,并通过公式WUE=Pn/Tr计算水分利用效率。测定时间为9:00—12:00;光合有效辐射为800 μmol·m-2·s-1,每个处理测3株,每株2~3片叶。叶绿素荧光参数测定:将不同控水时间的苗木放置于暗环境中12 h,采用Li-6400荧光叶室测定的初始荧光值和最大荧光值,并计算可变荧光值等数值。土壤含水率使用恒温烘干法,在温度105 ℃的烘箱内将采集土样进行烘考至恒重状态,称量土样干重并计算土壤含水率。计算公式:土壤含水率(%)=(土壤鲜重-土壤干重)/土壤鲜重SymboltB@
100。叶片相对含水量采用烘干称量法:将采集的叶片立即称取鲜重,将叶片置于蒸馏水中浸泡24 h,再称取叶片饱和鲜重,最后在85 ℃下烘干48 h至恒重,称取其干重。计算公式:叶片相对含水率(%)=(鲜重-干重)/(饱和鲜重-干重)100。丙二醛采用硫代巴比妥显色法提取(丁玉梅等,2013)。光合色素采用乙醇提取法:称取鲜叶0.2 g,用95%的乙醇定容于25 mL容量瓶中浸提,将溶液置于暗环境中24 h,最后在470、649、655 nm波长下测定的吸光值(柴胜丰等,2012)。
1.4 数据分析
采用SPSS 17.0统计分析软件对试验数据进行单因素方差分析(One-way ANOVA),并用最小显著差数法(LSD法)进行多重比较,用软件Excel 2007进行图标制作。
2 结果与分析
2.1 不同控水时间对叶片的光合参数及其水分利用率的影响
由图1可知,与CK相比,随着干旱时间的增加,其4个处理组的净光合速率分别平均下降了9.471%、30.617%、59.472%、89.648%。控水4 d其指数下降不明显,表明此阶段幼苗受到水分胁迫的影响不大;控水8 d其指数下降显著,说明东兴金花茶叶片的光合作用已经开始受到水分亏缺的影响;控水12 d其指数下降了59.472%,说明已经受到干旱胁迫的显著影响;16 d指数已经下降到89.648%,表明东兴金花茶光合作用已经遭到严重的抑制,严重的缺水对植物光合影响很大。随着控水时间增加,气孔导度分别下降了10%、34%、62%、80%,蒸腾速率分别下降了14.235%、36.588%、63.176%、82.824%。控水4 d,其叶片的气孔导度和蒸腾速率变化幅度都不大,无显著性差异;在控水8~12 d后,分别下降了30%~60%,说明水分亏缺造成叶片气孔导度及蒸腾速率的下降幅度显著,证明其叶片受到了较为严重的干旱胁迫,植株只能通过提高水分利用率来迅速适应其水分亏缺的状态;控水16 d后,叶片气孔导度和蒸腾速率都极显著下降了80%左右,说明严重的干旱缺水促使叶片气孔关闭,阻止叶片内部水分的过度蒸腾。
2.2 不同控水时间对叶片叶绿素荧光参数的影响
水分胁迫对植物的光合作用影响是多方面的,不仅影响光合电子传递和暗反应的有关酶系,还会引发植物光合系统的损伤(段仁燕等,2011)。利用叶绿体荧光动力学可高效检测水分胁迫对植物光合作用的影响。从表2不同胁迫时间的叶绿素荧光参数可以看出,随着控水时间不断增加,Fo下降趋势显著,由255下降至207左右,幼苗叶片的初始荧光值随着水分胁迫的加剧其受到了显著的影响,PS Ⅱ系统受到破坏,最大荧光值Fm参数未出现显著性差异。PS Ⅱ关闭后其荧光产量没有显著变化。Fv在控水0~12 d之间,其相对于CK并未有明显的变化,差异性不大,直到16 d其下降至782左右,降幅显著(P<0.05),说明叶绿体可变荧光值变低,原初电子受体QA的氧化态数量减少,使QA-QB传递电子的能力下降。Fv/Fm和Fv/Fo都随水胁迫天数的增加而不断下降,分别从0.806下降至0.754和4.170下降至3.080。光系统PS ⅡFv/Fm是衡量植物是否受到光抑制的敏感指标,正常情况下其参数稳定。东兴金花茶Fv/Fm正常值为0.806左右,低于正常值为光抑制现象,表明水分胁迫降低了PS Ⅱ原初光能转化效率,使PS Ⅱ潜在性活性受损,光合作用原初反应过程受到抑制。
2.3 不同控水时间对光合色素含量的影响
如表3所示,东兴金花茶叶片的光合色素中Chla、Chlb、Car随着控水时间的延长,其相对于CK,胁迫8 d内其含量都无显著性差异,胁迫至12 d以上其含量都出现了显著性下降。 Chla/Chlb比值可以直接反应集光复合体的捕光能力,较CK组其控水4 d的比值下降了0.12, 8~12 d又有逐渐上升的趋势,分别比控水4 d上升了0.074和0.224;控水16 d较控水12 d又下降了0.03。说明水分胁迫显著影响了东兴金花茶叶片光合色素的含量及比例;水分胁迫使其叶片的集光复合体捕光能力显著下降。Chla和Chlb的比值呈现降低又逐渐升高的趋势,这表明可能是其本身在水分胁迫状态下的一种光保护机制。
2.4 不同控水时间对土壤含水率的影响
由图2可知,控水时间0、4、8、12、16 d下的土壤含水率分别是26.527%、21.834%、15.065%、14.157%、11.447%。控水4 d(CL1)相对于CK土壤含水率下降了4.693%,干旱胁迫并不明显;CL2、CL3、CL4的土壤含水率下降显著,分别下降了11.462%、12.370%、15.180%。随着胁迫的时间不断延长,土壤含水率不断降低,说明水分的亏缺随着时间的延长逐渐严重,主要是由于土壤和叶片的水分不断被蒸发、蒸腾引起。
2.5 不同控水时间对叶片相对含水量的影响
从图3可以看出,在整个水分胁迫的控水过程中,东兴金花茶叶片的相对含水量呈现明显的下降趋势,叶片相对含水量分别是94.177%、90.282%、71.551%、62.620%、48.425%。与对照组相比,控水4 d 叶片相对含水量下降3.895%、8 d下降了22.626%、12 d下降了31.557%、16 d下降了45.752%。表明4 d的水控时间短,对植株幼苗葉片相对含水量的保水能力影响较小;8~12 d后,其叶片相对含水量下降显著,叶片的保水能力受蒸腾作用的影响下降显著,表明幼苗受到了轻、中度的干旱影响,叶片出现萎缩状态(图4);16 d后叶片相对含水量下降了几乎一半的水分,植株补给水分严重不足,叶片出现下垂萎缩和干枯现象,严重影响了植株的正常生长和发育状态。
2.6 不同控水时间对丙二醛含量的影响
丙二醛作为考察水分胁迫严重程度的指标之一,其含量的高低表明其细胞质膜结构和功能的损害程度(Qin et al., 2011)。由图5可知,相对于CK组(28.8 nmol·g-1),控水时间在8 d范围之内,其丙二醛含量变化不显著;控水12 d其丙二醛含量(30.4 nmol·g-1)小幅增加,说明其膜质发生过氧化反应,受水分胁迫程度不大;控水16 d,其丙二醛含量达到40.3 nmol·g-1,变化显著。以上结果表明幼苗受到干旱后其自身调节生理生化反应来适应干旱。若不及时补水,其细胞膜结构将受到严重损伤,甚至引起细胞死亡,植株停止生长。
3 讨论和结论
光合作用是绿色植物进行生长、发育所必不可少的条件,光合作用可将太阳能转化为植物生长发育所需的化学能,储存在有机物化合物中,为其生长发育提供能量(王学文等,2010)。但是,光合作用经常会受外界环境因子的干扰而发生变化,例如在干旱、半干旱、水涝、光照强度等因素都会造成光合效率的改变(丁玉梅等,2013)。水分是环境中最重要的生态因子,水分亏缺时,会引发气孔关闭、酶活性及光系统活性下降,造成光合作用减弱并抑制植物的生长(王继玥等,2017)。本研究结果表明干旱胁迫造成光合速率下降的原因主要分为气孔限制因素和非气孔限制因素,前者主要是在轻度胁迫时干旱胁迫使气孔关闭而导致气孔导度、胞间 CO2浓度、蒸腾速率下降,光合作用随之下降,后者是在严重胁迫时,导致光合色素解体、RuBP羧化酶活性、PS Ⅱ结构受损等非气孔因素(申长青,2016)。在CL1中,干旱胁迫初期,气孔导度的下降与光合速率降低相对一致,继续干旱胁迫,CL2的气孔导度的下降幅度比光合速率大,持续干旱12 d后,光合速率的下降幅度较气孔导度大,这说明东兴金花茶幼苗在干旱初期光合速率下降主要受气孔关闭的影响,植物通过提高水分利用率和降低蒸腾速率来适应干旱环境。随着干旱胁迫的加剧,气孔导度的下降幅度比光合速率的下降幅度大,说明此阶段是气孔限制和非气孔限制交替或综合调节所致(李杰芬,1980);当光合速率的下降幅度超过气孔导度的下降幅度,表明此阶段干旱严重,东兴金花茶叶片的PSⅡ的结构和光能转换效率受到损伤与破坏,光合效率下降主要是非气孔限制因素造成。
光合色素是植物光合作用的物质基础,其参与吸收、传递光能或引起原初光化学反应。不同环境状况的改变就会引起光合色素的变化(吴甘霖等,2010),光合色素含量的高低很大程度上可以反映植物在不同干旱胁迫条件下的生长状况和叶片的光合能力(应叶青等,2009)。本研究结果显示东兴金花茶光合色素的含量低,说明了金花茶对干旱敏感、不耐旱的特征。
叶片相对含水量是植物耐旱性的重要指标,下降幅度小速率慢,说明其在此状态下叶片保水能力强、抗旱性强(陈少瑜等,2004;陈荣建等,2017)。本研究结果表明东兴金花茶幼苗在控水4 d的情况下,叶片相对含水量下降幅度小,说明具有一定的耐旱性。植物叶片的含水量能准确地反映植物在水分胁迫条件下其体内水分的真实亏缺情况,而且与植物幼苗外观特征保持相对一致。东兴金花茶幼苗叶片在水分充足时,叶光泽油绿、舒展、长势很强;水分不足时,叶片萎蔫、叶色变浅、长势很弱;水分亏缺严重时,叶片开始打卷。证明东兴金花茶的光合作用对干旱胁迫敏感。
土壤含水率对植物吸收水分、保持自身生长和发育至关重要。干旱程度的加剧严重影响了土壤含水率,直接影响着东兴金花茶的光合作用及其生理生化代谢反应。本研究中,东兴金花茶幼苗在控水4 d情况下,土壤含水率为21.834%,下降不显著;控水8~12 d,东兴金花茶的光合指标下降显著,土壤含水率下降至14.157%~15.065%,其叶片萎蔫、打卷,而低于此水平东兴金花茶幼苗光合作用受到了严重的抑制。植株因过度干旱无法正常生存而死亡,表明东兴金花茶幼苗对水分胁迫的耐受极限土壤含水率为14.157%~15.065%,比金花茶幼苗对干旱胁迫的耐受极限15.04%~16.54%(柴胜丰等,2015)稍低些,说明东兴金花茶幼苗比金花茶幼苗对干旱的耐受能力强,这进一步证实了金花茶组植物对干旱胁迫十分敏感。
综上所述,东兴金花茶幼苗控水时间在4 d情况下东兴金花茶可以提高自身水分利用效率来抵抗干旱,说明东兴金花茶幼苗对水分胁迫具有一定的适应能力和响应机制。东兴金花茶幼苗对水分胁迫的耐受极限土壤含水率为14.157%~15.065%,说明东兴金花茶幼苗对干旱胁迫较为敏感,这与东兴金花茶的生境相符,其多分布于荫蔽的林下和潮湿的沟谷两旁,表明东兴金花茶对干旱胁迫的敏感、不耐干旱的生态特性可能是限制其种群扩散的一个重要原因。
参考文献:
AN DS, 2011. Chlorophyll fluorescence based water stress diagnosis for cut Lilium grown in greenhouse[D]. Nanjing:Nanjing Agricultural University.[安东升, 2011. 温室切花百合对干旱胁迫响应的叶绿素荧光诊断研究[D]. 南京:南京农业大学.]
CHAI SF, TANG JM, WANG ML, et al., 2015. Photosynthetic and physiological characteristics of Camellia petelotii seedlings under drought stress[J]. Acta Bot Boreal-Occident Sin, 35(2):322-328.[柴胜丰, 唐健民, 王满莲, 等, 2015. 干旱胁迫对金花茶幼苗光合生理特性的影响[J]. 西北植物学报, 35(2):322-328.]
CHAI SF, WEI X, SHI YC, et al., 2012 . Effect of strong light stress on the growth, biomass and chlorophyll fluorescence parameters in seedlings of endangered plant Camellia nitidissima[J]. Bull Bot Res, 32(2):159-164.[柴勝丰, 韦霄, 史艳财, 等, 2012. 强光胁迫对濒危植物金花茶幼苗生长和叶绿素荧光参数的影响[J]. 植物研究, 32(2): 159-164.]
CHEN RJ, OU J, WANG LJ, et al., 2017. Effects of inoculated different ericoid mycorrhizas strains on physiological characteristics of Rhododendron annae under water stress[J]. J Cent S Univ For & Technol, 37(9):43-48.[陈荣建, 欧静, 王丽娟, 等, 2017. 水分胁迫下接种不同 ERM 菌株对桃叶杜鹃幼苗生理特性的影响[J]. 中南林业科技大学学报, 37(9):43-48.]
CHEN SY, LANG NJ, LI JY, et al., 2004. Changes of leaf relative water content, relative plasma membrane permeability and proline content of seedlings of three species under drought stress[J]. J W Chin For Sci, 33(3):30-33.[陈少瑜, 郎南军, 李吉跃, 等, 2004. 干旱胁迫下3树种苗木叶片相对含水量、质膜相对透性和脯氨酸含量的变化[J]. 西部林业科学, 33(3):30-33.]
DENG Y, WANG B, SU WH, et al., 2008. Adaptability of endangered plant Dipteronia dyerana under irradiance and irradiance and water stress[J]. Chin J Ecol, 27(6):888-893.[邓云, 王冰, 苏文华, 等, 2008. 濒危植物云南金钱槭对光、水环境的适应性[J]. 生态学杂志, 27(6):888-893.]
DING YM, MA LH, ZHOU XG, 2013. Effects of drought stress on free proline and malonaldedyde contents in potato leaves and correlation analysis of drought-tolerant level among different varieties[J]. SW Chin J Agric Sci, 26(1):106-110.[丁玉梅, 马龙海, 周晓罡, 2013. 干旱胁迫下马铃薯叶片脯氨酸、丙二醛含量变化及与耐旱性的相关性分析[J]. 西南农业学报, 26(1):106-110.]
DUAN RY, WU GL, WANG ZG, et al., 2011. Effects of temperature stress on chlorophyll fluorescence parameters of Fragaria ananassa Duch cv. Toyonoka[J]. Chin J Spectrosc Lab, 28(5):2249-2252.[段仁燕, 吴甘霖, 王志高, 等, 2011. 温度胁迫对草莓叶绿素荧光参数的影响[J]. 光谱实验室, 28(5):2249-2252.]
FU QS, LI HL, CUI J, et al., 2009. Effects of water stress on photosynthesis and associated physiological characters of Capsicum annuum L.[J]. Sci Agric Sin, 42(5):1859-1866.[付秋實, 李红岭, 崔健, 等, 2009. 水分胁迫对辣椒光合作用及相关生理特性的影响[J]. 中国农业科学, 42(5): 1859-1866.]
HU XJ, LIU YC, PEI Y, et al., 2020. Effects of soil water stress on physiological characteristics of Lilium brownii seedlings[J]. J Henan Agric Sci, 49(1):111-117.[胡小京, 刘玉彩, 裴芸, 等, 2020. 水分胁迫对野百合幼苗生理特性的影响[J]. 河南农业科学, 49(1):111-117.]
HUANG SX, TANG WX, WANG Y, et al., 2006. Distributional characteristics of rare and endangered plants in Guangxi[J]. J Fujian For Sci Technol, 33(4):169-173.[黄仕训, 唐文秀, 王燕, 等, 2006. 广西珍稀濒危保护植物分布特征[J]. 福建林业科技, 33(4):169-173.]
JIANG YS, TANG H, WEI X, et al., 2010. Introduction and acclimatization of Camellia tunghiensis a rare and endengered plant[J]. Guihaia, 30(3):362-366.[蒋运生, 唐辉, 韦霄, 等, 2010. 濒危珍稀植物东兴金花茶的引种驯化的研究[J]. 广西植物, 30(3):362-366.]
LI JF, 1980. The relationship between photoreaction and dark reaction in photosynthesis[J]. Bull Biol, 2:21-22.[李杰芬, 1980. 光合作用中光反应和暗反应的关系[J]. 生物学通报, 2:21-22.]
LIU L, 1999. Rare ornamental plants in the world—Camellia tunghinensis[J]. Guangxi For Sci, 28(2):94-96.[刘林, 1999. 世界珍稀观赏植物——金花茶[J]. 广西林业科学, 28(2):94-96.]
QIN YR, NONG YC, HUANG JB, et al., 2011. The comparison of contents of malondialdehyde and proline in the area of karst area in north-weastern Guangxi Province[J]. Agric Sci Technol, 12(4):469-473.
SHEN CQ, 2016. The study on adaptation of droughty and shade stress of Loropetalum subcordatum [D]. Guangzhou:South China Agricultural University.[申长青, 2016. 濒危植物四药门花对光照和水分胁迫的适应性研究[D]. 广州:华南农业大学.]
TANG JM, CHEN ZY, WEI X, et al., 2014. Optimization of SSR-PCR reaction system and primer screening of Camellia tunghinensis [J]. Genom Appl Biol, 33(2):398-404.[唐健民, 陈宗游, 韦霄, 等, 2014.东兴金花茶 SSR-PCR 反应体系的优化及引物筛选[J]. 基因组学与应用生物学, 33(2):398-404.]
WANG ML, TANG H, HAN Y, et al., 2017. Effects of water stress and rewatering on eco-physiological characteristics of Illicicum difengpi[J]. Guihaia, 37(6):716-722.[王满莲, 唐辉, 韩愈, 等, 2017. 水分胁迫与复水对地枫皮生理生态特性的影响[J]. 广西植物, 37(6):716-722.]
WANG JY, SHI DH, DU B, et al., 2017. Effect of water stress on the physiological and photosynthesis characteristic of Okra (Abloschus esculentus L.)[J]. Chin J Trop Crop, 38(9):1614-1619.[王继玥, 石登红, 杜斌, 等, 2017. 水分胁迫对黄秋葵生理以及光合特性的影响[J]. 热带作物学报, 38(9):1614-1619.]
WANG XW, FU QS, WANG YJ, et al., 2010. Effects of water stress on growth and photosynthetic system characters of Lycopersicon esculentum L.[J]. J Chin Agric Univ, 15(1):7-13.[王學文, 付秋实, 王玉珏, 等, 2010. 水分胁迫对番茄生长及光合系统结构性能的影响[J]. 中国农业大学学报, 15(1):7-13.]
WU FQ, ZHANG XJ, SHEN SK, et al., 2015. Effect of soil nutrition and water content on the seedling growth and physiological characteristics of Euryodendron excelsum H. T. Chang.[J]. Guangdong Agric Sci, 20:45-51.[吴富勤, 张新军, 申仕康, 等, 2015. 土壤养分与水分对猪血木幼苗生长及生理特性的影响[J]. 广东农业科学, 20:45-51.]
WU GL, DUAN RY, WANG ZG, et al., 2010. Effects of drought stress and rehydration on chlorophyll fluorescence characteristics in Fragaria ananassa Duch.[J]. Acta Ecol Sin, 30(14):3941-3946.[吴甘霖, 段仁燕, 王志高, 等, 2010. 干旱和复水对草莓叶片叶绿素荧光特性的影响[J]. 生态学报, 30(14):3941-3946.]
YANG QH, LI XQ, YANG H, et al., 2010. Photosynthetic ecophysiological characteristics of Camellia nitidissima seedlings[J]. J Beijing For Univ, 32(2):57-63.[杨期和, 李旭群, 杨和, 等, 2010. 金花茶幼苗光合生理生态特性研究[J]. 北京林业大学学报, 32(2):57-63.]
YANG QG, SONG HT, ZHANG H, 2012. Effects of different substrates on seedling development of Camellia tunghinensis Chang[J]. J Green Sci Technol, 12:107-108.[杨泉光, 宋洪涛, 张洪, 2012. 不同基质对东兴金花茶种苗发育的影响[J]. 绿色科技, 12:107-108.]
YING YQ, GUO J, WEI JF, et al., 2009. Photosynthetic andchlorophyll fluorescent responses of Phyllostachys pubescens seedlings to water deficiency stress[J]. J Beijing For Univ, 31(6):128-133.[应叶青, 郭璟, 魏建芬, 等, 2009. 水分胁迫下毛竹幼苗光合及叶绿素荧光特性的响应[J]. 北京林业大学学报, 31(6):128-133.]
ZHAO LL, XIN B, LIU AQ, 2019. Effects of different water and fertilizer control treatments on tomato fruit development and flavor quality in solar greenhouse[J]. Liaoning Agric Sci, (5):22-25.[赵丽丽, 辛彬, 刘爱群, 2019. 不同控水控肥处理对日光温室番茄生长发育及果实品质的影响[J]. 辽宁农业科学, (5):22-25.]
(责任编辑 周翠鸣)
